はじめに
高等学校の生物IBの遺伝の分野では組換えを利用した遺伝地図の作成法の原理が扱われている。教科書に書かれている範囲では、それほど理解困難であるとは思えないが、入試対策などで一歩踏み込んだ内容になると、教科書の説明は明らかに不十分であり、正しく理解することは困難である。Jflyのウェブページに高校の遺伝教育にみられる問題点の一つとして、遺伝地図の解釈の間違いを紹介したところ、何人かの高校の先生方からこれに関連した質問を受けたが、その内容からみて、この問題に関する誤解は想像以上に深刻であるらしいことがうかがえた。質問を寄せられた先生方は、いずれも研究熱心な方々であり、自分がきちんと理解してから生徒に教えたいという熱意が感じられた。しかしながら、古典遺伝学の最も基本的な事柄の一つである乗換えや組換えについてきちんと説明した日本語の参考書や教科書がほとんどみられないこともあり、自己流の解釈や怪しげな解説に頼ることになっているらしい。ここでは、最近見聞きした典型的な誤解の例を取り上げ、解説することにしたい。
乗換えと組換え
本題に入る前に、基本的な用語である乗換えと組換えの違いを明確にしておきたい。目にとまった範囲の教科書の記述をみるかぎり、この2つの用語を混同しているような例はみられない。しかし、先生方からの質問の中では、明らかに混同しているとしか思えないケースも少なくない。
乗換えというのは、第一減数分裂において、相同染色体が互いの腕の一部を交換するという現象である。DNA分子のレベルでみると、まだ未解明の点も多いきわめて複雑な過程であるが、ここでは結果だけの理解で充分であろう。結果から見ると、2本の相同染色体が同一個所で切断され、異なった染色体つなぎ合わされるという物理的な現象が乗換えである。一方、組換えは、配偶子(精子や卵子など)の持つ特定の染色体上の遺伝子の組み合わせが両親から受け継いだ2本の染色体のいずれとも異なるという遺伝学的な現象のことをいう。つまり、乗換えと組換えは、互いに無関係ではないが、全く別次元の現象であることをしっかり認識することが大切である。
組換えはいうまでもなく乗換えがその原因である。したがって、注目した2つの遺伝子座の間で組換えが観察された場合、この区間で少なくとも1回の乗換えが起こったことを示す。しかし、逆は必ずしも真ではない。組換えが見られない場合、乗換えがないとはいえないのである。これは、複数回の乗換えが起こると、2つの遺伝子座の組み合わせが元に戻ってしまい、組換え体として観察されない場合があるためである。
組換えによる遺伝地図作成の原理
モーガンの弟子の一人であるスターテバント(Sturtevant)は、乗換えとその結果である組換えを利用すると遺伝子座間の距離を測定でき、これにもとづいて染色体上の遺伝子座の位置を示す地図(遺伝地図)を作成できることに気付いた。乗換えが染色体のどこでも一様に起こると仮定すれば、注目した2つの遺伝子座の間で乗換えの起こる確率は、遺伝子座間の物理的な距離に比例するはずで、したがって、乗換えの頻度がわかればそれから相対的な距離が推定できるというのがその原理である。
しかし、乗換えそのものを直接観察することは通常不可能である。そこで、乗換えの結果である組換えの頻度を利用しようとするのである。当初、乗換えは減数分裂の染色体の複製前に2本の相同染色体の間で起こるものと想定されたため、二重乗換えの確率が無視できるような近い距離にある遺伝子座の間の組換えの頻度(組換え価)は乗換えの確率に等しいと考えられていた。すなわち、組換え価=仮想的な乗換え価とみなして組換え価の値そのものを距離の単位として用い、1%の頻度で組換え型配偶子を生ずる距離を1センチモーガン(1cM)と定義したのである。したがって、地図距離を示す数値そのものは組換え価から求めるのであるが、その意味するところは乗換えの頻度の推定値なのである。
遺伝地図が組換え価にもとづいて作られることから、地図距離は組換え価に相当すると誤解している人をかなり見受ける。例えば、50cM離れた遺伝子座間の組換え価は50%だろうといったケースである。後で述べるように、この場合の組換え価は実際には40%を越えないのである。
その後、乗換えは染色体の複製後に起こることがショウジョウバエやアカパンカビなどを用いた研究によって明らかになり、その結果、地図距離は後述するように乗換え頻度の1/2に相当することになったため、乗換え価という用語はとかく混乱を招きやすいことから、最近はほとんど用いられなくなった。
組換え価が50%を越えない理由
組換え価が50%を越えないことは、学習参考書などにも記述されていることが多い。しかし、このことを教科書や参考書などでよく目にするように、2本の相同染色体の間で乗換えが起こるかのように説明した図などから理解することは困難であろう。そのためか、奇妙な解説が流布されているらしい。何人かの高校の先生から耳にした説を一つ紹介する。
それは、2つの遺伝子座の間で乗換えの起こる確率をpとすると、組換え価は、p/(1+p)で表せるので、p=1(つまり100%の確率)のとき、組換え価は1/2になり、これを越えることはないというものである。このような説を誰が言い出したのかわからないので、直接説明を聞いたわけではないが、結論から想像するに、以下のような計算によるらしい。
組換え価をrとすると、p<1なので、下記の等比級数は収束し、
r=p - p2 + p3 - p4 + ....= p/(1+p)
この式の各項は、それぞれ乗換えが1回、2回、3回・・・起こる確率である。また、p2やp4の係数がマイナスになっているのは、偶数回の乗換えが起こると組換え体として観察されないために、これを差し引く必要があると考えたのであろう。一見したところもっともらしく見えるし、数学的には上式に誤りはない。しかし、確率の計算としては完全な間違いである。このことを簡単に確かめるには、1回以上乗換えの起こる確率を全て足し合わせてみればよい。
p + p2 + p3 +....= p/(1-P)
となり、p=0.5のとき1となり、これよりpが大きいと1以上になってしまい、乗換えが起こる確率が1を越えるという奇妙な結果になる。また、乗換えが起こらないというケースも当然あるにも関わらず、この計算には含まれていないのである。
組換え価が50%を越えないのは、実は乗換えが染色体の複製後に、4本の染色分体のうち2本の非姉妹染色分体の間で起こるからである。このことは、最近の教科書の減数分裂を示した図にはちゃんと書かれているものが多いにも関わらず、組換えや遺伝地図作成の説明になると染色体が2本しか書かれていないことも誤解が生ずる原因の一つであろう。
まず、注目した2つの遺伝子座aとbの間で1回の乗換えが起こる場合について考えることにする。図1に示すように、それぞれの遺伝子座には対立遺伝子aとAおよびbとBがあるものとし、遺伝子型はa b/A B とする。乗換えは、4本の染色分体のうち任意の2本の非姉妹染色分体間で起こるものとする。姉妹染色分体間でも起こる可能性はあるが、実はこれはよくわかっていない。しかし、姉妹染色分体は遺伝的に全く等しいので、仮に乗換えが起きても何の変化も見られないので、ないものとして無視しても差し支えない。姉妹染色分体は実際には区別不能であるが、説明の便宜上、ABの遺伝子を持つ染色分体を1と1’、また、a bの遺伝子を持つ染色分体を2と2’のように表すことにする。いま、乗換えが1’と2の間で起こると、Abと a Bの組換え型染色体が2本でき、1と2’は非組換え型となるので、4本の染色分体のうち組換え型は2本となり、組換え価は50%になることは容易に理解できよう。
このように説明すると、一つの減数分裂で4種類の配偶子が作られるものと考える人が少なくないが、必ずしもそうではない。例えば、ショウジョウバエでは、乗換えは雌でしか起こらないが、卵核になるのは減数分裂でできた4個の核のうちの一つだけであり、残りの3つは極体に捨てられてしまう。すなわち、1個の卵について組換えをみると、0%か100%の2通りしかないのである。0%のものと100%のものが1:1の割合で生ずるので、多数の卵を調べると平均50%になることに注意してほしい。
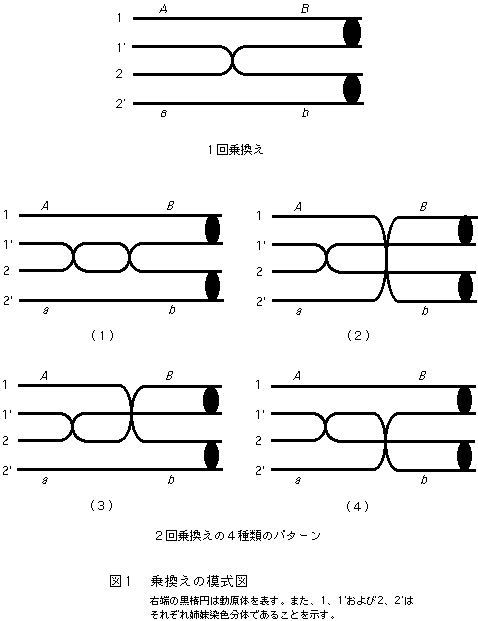
次に、2回の乗換え(二重乗換え)の場合について考えることにしよう。まず、1回目の乗換えが1’と2の間で起こり、2回目も同じく1’と2の間で起きたとすると、遺伝子の組み合わせは元に戻ってしまうので、組換え型染色体の割合は0になってしまう(図1の(1))。このような組み合わせは、このほかに1-2、1-2’、1’-2’の3通りあるので、つごう4通りの組み合わせになる。次に、2回目の乗換えが1回目の乗換えに参加しなかった染色分体の間で起こる場合を考える。1回目が1’-2、2回目が1-2’間といったケースである。この場合、減数分裂で生ずる4本の染色体の全てが組換え型になるので、組換え価は100%になる(図1の(2))。このような組み合わせも4通りあることは説明するまでもないであろう。もう一つの可能性は、2回目の乗換えが1回目の乗換えに関与した染色分体と関与しなかったものとの間で起こるケースである(図1の(3)と(4))。例えば、1回目が1’-2、2回目が1-2というような場合である。これによって生ずる4本の染色体のうち、1と2の2本は組換え型、1’は二重乗換えのために非組換え型になり、2’は乗換えに関与しないので当然非乗換え型である。結局、組換え型の割合は50%となる。このような可能性は、1回目の乗換えの4通りの可能性のそれぞれについて、2回目の乗換えに関与する染色体は2通りずつあるので、つごう8通りの組み合わせがある。
二重乗換えの結果をまとめてみると、組換え価0%、50%、100%の可能性が4:8:4(つまり、1/4:2/4:1/4)の割合でみられることになり、平均組換え価は、
0/4+1/4+1/4=2/4=50%
になる。
三重乗換え(3回乗換え)の場合は、可能性が64通りにもなるため、図に示すと煩雑になるので、別の考え方で説明することにする。いま3回の乗換えの結果生じた任意の1本の染色体に注目すると、3箇所の乗換えサイトについて、それぞれ乗換えが起こらなかった場合(Nで表すことにする)と起きた場合(Yで表す)の2通りの状態がある。姉妹染色分体のどちらが乗換えに関与するかはランダムなので、あるサイトがNになるかYになるかはコインを投げたときに表が出るか裏がでるかと同じで、確率は等しく、1/2になる。3箇所の乗換えサイトのどこでも乗換えを起こさなかった場合を、NNNと表すことにすると、このような染色体は非組換え型になることはいうまでもない。また、3箇所とも乗換えを起こした場合、つまりYYYは、乗換え回数が奇数なので組換え型になる。同様にして、3回乗換えの結果生ずる染色体のタイプとその出現確率および組換え確率は表1にまとめた。
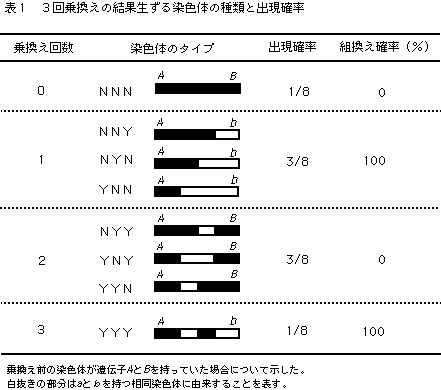
これから平均組換え価(RV)は、
RV=0x1/8+100x3/8+0x3/8+100x1/8=400/8=50(%)
それぞれのタイプの出現確率が(N/2+Y/2)3を展開したものであることに注意すると、任意の回数nのケースに拡張できることが容易にわかるであろう。すなわち、n回の乗換えが起きた場合、任意の1本の染色体で0,1,3,...x回の乗換えが確率P(x)は、(N/2+Y/2)nを展開したときの係数になり、
P(x) = nCx(1/2)x(1/2)n-x = (1/2)n n!/(n-x)!x!
これから、例えば4回乗換えの場合は、それぞれ1/16:4/16:6/16:4/16:1/16の確率で出現することがわかる。このような確率分布を二項分布というが、これは左右対称な分布なので、組換え価0%の染色体の出現確率と100%の出現確率が等しくなる。つまり、平均組換え価は、乗換えが何回起ころうとも常に50%になるのである。
乗換えと組換えの関係
注目した2つの遺伝子座の間で乗換えがランダムに起こるものと仮定すると、この区間内で0,1,2,3,...x回の乗換えが起こる確率P(x)はポアソン分布に従うと考えられるので、平均乗換え回数をmとすると、
P(x) = e-mmx/x!
これから、乗換えが全く起こらない確率は、
P(0) = e-mm0/0! = e-m
となる。乗換えが1回以上起こる確率はこの余事象なので、
P(x>0) = 1-e-m
さて、乗換えが1回以上おこると組換え価が50%になることは上でみたとおりなの で、組換え価をrとすると、
r = (1-e-m)/2 となる。
平均1回の乗換えが起こるような距離にある2つの遺伝子座間に見られる組換え価は、上式でm=1とおくと、r=0.316(31.6%)と予想される。先に述べたように、地図距離は乗換えの頻度を反映するので、平均1回乗換えの起こる距離は50cM(=0.5M)に相当する。つまり、50cM離れた遺伝子座間で観察される組換え価は、乗換えがランダムに起こると仮定すると、32%にしかならないのである。
モーガン単位で表した遺伝距離(dとする)と乗換え頻度(m)との間には、m=2dの関係があるので、上式は、
r = (1-e-2d)/2 と表すことができ、これから、
d = -ln(1-2r)/2 (lnは自然対数を示す)
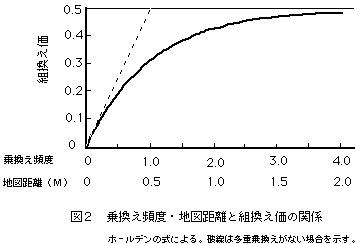
この関係を利用すると、実験で観察された組換え価から地図距離を推定できる。このような式は地図関数と呼ばれるが、これはその一つであり、ホールデン(Haldane)の地図関数といわれるものである。遠距離にある遺伝子座を用いて遺伝地図を作成する必要のある場合には、このような地図関数を使って組換え価を補正し、地図距離の推定値とする。
図2には乗換え頻度・地図距離と組換え価の関係をホールデンの式にもとづいて示した。地図距離が小さい範囲では地図距離と組換え価はほぼ直線的な関係にあるため、組換え価はほぼ正確に地図距離を反映するが、距離が離れるにしたがって直線からずれるため、組換え価は地図距離の過小推定値になる。そこで、地図関数により補正することが必要になるが、組換え価が50%に近くなると補正したとしても誤差が大きくなり実用には耐えない。
ホールデンの式は乗換えがランダムに起こることを仮定して導かれた。しかし、実際には乗換えはランダムではない。ある個所で乗換えが起こると、その近傍では乗換えが起こりにくくなる干渉と呼ばれる現象が一般的に見られるためである。干渉がどのような機構によるかはまだ明かではないので、詳しい説明は省略するが、経験的には10cM以下の距離にある遺伝子座の間では多重乗換えは事実上見られないことが知られている。このため、より正確な地図関数を作るには干渉を考慮に入れる必要があるが、干渉の強さは染色体の場所によっても変わるため、完ぺきといえるような地図関数は今のところ存在しない。干渉が距離に反比例するという比較的単純な仮定にもとづいた地図関数として、コサンビ(Kosambi)の式があり、広く用いられている。これによると、組換え価rと地図距離dの関係は、
r = (tanh2d)/2 (tanhは双曲線関数)
d = ln{(1+2r)/(1-2r)}/4
この式で50cM離れた遺伝子座間の組換え価を求めると、38.1%となり、ホールデンの式による場合よりは大きくなる。これはいうまでもなく多重乗換えの頻度が干渉のために減少することによるが、現実に観察される組換え価もこれに近い。
最後に、蛇足であるが、組換え価が50%を越えないという記述を読んで、実際の実験でも50%を越える組換え価は決して観察されることはないと受け取る人を見受けた。組換えは確率現象であり、当然誤差を伴うので、50%を越える組換え価が観察されることもある。組換え価は二項分布と呼ばれる確率分布に従うと考えられるので、その分散は、r(1-r)/n(nは標本数)、また、標準偏差はこの平方根になる。例えば、真の組換え価が45%とし、100個の配偶子から組換え価を求めたとすると、標準偏差は約5%になる。したがって、観察される組換え価が50%以上になる確率は15%を越えるのである。
おわりに
以上に解説したような内容について、確率や統計学を学習していない高校生に理解させることはほとんど不可能であろう。また、その必要もないと考える。とかく誤解の多い組換え価が50%を越えないことも、それ自体は至極当たり前のことであり、とりたてて教えるまでもないであろう。しかし、組換えを利用した遺伝地図作成の原理について授業で扱う場合、センチモーガン単位で表した地図距離が乗換えの頻度を反映したものであり、組換え価と1:1に対応するものでないことは最低限理解させることが必要であると思う。この点で、二重乗換えの可能性を無視して説明した教科書や学習参考書の多いことは困った問題である。
減数分裂における乗換えとそれによる組換えは有性生殖の根幹をなすものであり、生物学的にはきわめて重要な現象なので、少なくとも教える先生方には正しく理解して頂きたいと願っている。本稿がその一助となれば幸いである。
|