<- Previous Issue | Next Issue ->
Drosophila Newsletter (Japanese) No. 17
The Newsletter of the Japanese Drosophila Research Conference
(in Japanese)
July 1999
ショウジョウバエ通信 No.17
1999年 7月
|  |
← 前号 | 次号 →
この通信は研究室単位でお送りしています。研究室の方全員に読んでいただけるようご配慮お願いいたします。
第4回研究集会の御案内(第4報)
第4回研究集会準備委員会 西田 育巧
(名古屋大学大学院理学研究科生命理学専攻)
いよいよ研究集会も間近となりました。多数の方の参加・演題申し込みを頂き、有難うございました。プログラムがほぼ確定いたしましたので、お知らせいたします。暑い最中(特に名古屋の夏は暑い!)ではありますが、この日に向けて体力を調整されて、多数の方のお出でをお待ちいたしております。なお、うっかりしていて、参加登録を忘れておられる方も、当日参加大歓迎です。振るって御参加下さい。
1.日時
1998年8月2日(月)13:00 ~4日(水)15:00
(受付開始:8月2日 12:00から)
2.場所
愛知県がんセンター国際医学交流センター
(〒464-8681 名古屋市千種区鹿子殿1ー1)
会期中の緊急連絡先(FAXのみ): 052-763-5233
愛知県がんセンター研究所生物学部気付
3.交通
会場への交通の便があまり良くありませんが、御了承願います。
◎名古屋駅より
1)地下鉄東山線「本山」駅下車、本山より市バス(光が丘行、猪高車庫行のいずれでもよい)で「自由が丘」にて下車(全所要時間約30分)。または、本山よりタクシー(約1000円)。バス停から、進行方向右側に愛知県がんセンターがあります(徒歩3分)。
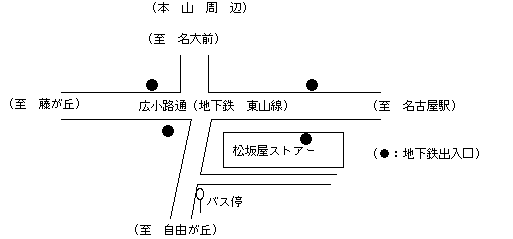
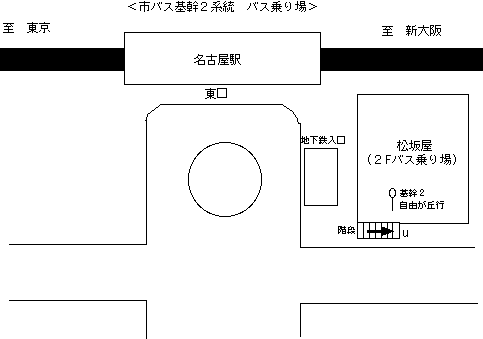
2)市バス基幹2系統、自由が丘行で終点「自由が丘」にて下車(所要時間約45分)。
◎名古屋空港より
「名古屋駅」行または「栄」行空港バスで「黒川」にて下車(最初のバス停)、地下鉄名城線に乗換え、「栄」でさらに東山線に乗換えて、「本山」下車。本山より市バス(光が丘行、猪高車庫行のいずれでもよい)で「自由が丘」にて下車。
4.宿泊
宿泊の予約は、各自でお願いします。通信16号でお知らせしましたように富士ツーリストに、割安のホテルを斡旋してもらっております。一応、申し込み閉め切りは7月9日(金)となっておりますが、部屋に余裕のある限り、できるだけ直前まで対応して頂くようにお願いしております。直接、下記の旅行社にお問い合わせ下さい。
(株)富士ツーリスト
〒460-0008 名古屋市中区栄3-32-26 六合ビル4F
TEL (052) 261-4621 FAX (052) 251-6913
担当:富田 千草・宇野 せき子
5.受付
受付でチェックをして、名札カードと要旨集を受け取って下さい。名札カードには氏名・所属を各自で御記入願います。参加費未納の方、および当日参加申込の方は、下記の当日参加費を申し受けます。
当日参加費 有給者(含PD、研究員) 5000円
院生・学生・研究生 3500円
受付の際にできるだけ、懇親会への参加申込も同時に行うようにお願いいたします。要旨集が余分に必要な方には、1000円(ほぼ実費)で販売の予定です。
6.会場
会場は、メインホール、大会議室、視聴覚室の3会議場からなっております。会場内は、全面的に禁煙になっておりますので、御協力の程お願いいたします。また、メインホールでの飲食は、いっさい認められておりませんので、宜しくお願いいたします。昼食の弁当の販売を予定しておりますが、食事は大会議室およびロビーにてお願いいたします。メインホールでは、食事をしないようにお願いします。
講演および総会などはメインホールで行います。ポスターパネルおよび休憩所は、大会議室と廊下に設定します。視聴覚室は、クローク(セルフサービス)として使用します。
7.ポスター
下記のポスター一覧で、各自のポスター番号を御確認下さい(要旨は、近日中にショウジョウバエ研究会のホームページから閲覧可能)。
ポスター掲示用のパネルの大きさは、横90cm×縦210cmです(下50cm位は使わない方が良い)。これに収まるように御準備下さい。
ポスターパネルは、大会議室と廊下に並べますので、指定の番号のところに掲示して下さい。8月2日(月)12:00から掲示可能です。期間中は、ずっと掲示することができます。ポスターの撤去は、8月4日(水)12:45までにお願いします。13:00より、業者がポスターパネルの撤去に取りかかります。
ポスターの説明を8月3日(火)午前中に行います。ポスター番号が奇数の方は、10:00~11:00、偶数の方は11:00~12:00の間、ポスターの前で、説明をお願いします。
8.スライド
シンポジウム・講演・テクニカルセミナー・討論会でのスピーカーの方は、メインホール前のスライド受付で、時間的余裕をもって、スライドの受付をお願いします。ビデオ映写も可能ですが、ビデオ使用の場合は、前もって、事務局(西田)まで、ご連絡下さい。
9.総会
8月3日(火)のシンポジウム終了後、引き続いて総会を開催しますので、御出席願います。次期世話人・第6回研究集会代表者の選出・承認、事務局からの報告、その他を行いますので、必ず御出席下さい。
10.懇親会
3日(火)の総会終了後、18:00より、名古屋大学シンポジオン内(豊田講堂の裏)のレストラン「ユニバーサルクラブ」にて、懇親会を開きます。振るって御参加下さい。参加申し込みは、できるだけ受付時にお願いします。
会場へは、市バス1本で、「自由が丘」から「名古屋大学前」まで、行くことができます(所要時間約30分)。また、病院玄関前のタクシー乗り場から、タクシーの乗り合いで行かれるのをお勧めします。
懇親会参加費 有給者(含PD、研究員) 4000円
院生・学生・研究生 2500円
11.昼食
愛知がんセンター病院内食堂と、がんセンター周辺に若干の食堂があります。また、昼食には弁当の販売を計画しています(予約制)ので、御利用下さい。詳しい案内は、当日会場にて行います。
食事は、大会議室または、廊下でお願いします。メインホールで食事はできません。
12.その他
・ロビーにて飲み物を用意する予定です。
・クローク(視聴覚室)には、各自で荷物を置いて下さい。不審者の出入りには注意をする予定ですが、貴重品などの管理は各自でお願いいたします。
・運営委員は、カラーの名札を着用しておりますので、何かありましたら、お問い合わせ・御相談下さい。
それでは、皆様のお越しをお待ちしております。
第4回研究集会代表者
西田 育巧
TEL: 052-789-2472
FAX: 052-789-2511
e-mail: nishida@bio.nagoya-u.ac.jp
第4回研究集会プログラム
8月2日(月)
12:00 受付開始
ポスター掲示開始
13:00~13:05 開会
13:05~16:30 講演
<座長:井上 喜博(愛知がんセ研)>
L-1 13:05 倉田 祥一朗(東北大・薬)
L-2 13:35 多羽田 哲也(東大・分生研)
L-3 14:05 広海 健(遺伝研・発生遺伝)
(14:35~15:00 休憩)
<座長:中村 真(基生研・形態形成)>
L-4 15:00 三浦 正幸(阪大・医)
L-5 15:30 山口 政光(愛知がんセ研・生物)
L-6 16:00 布山 喜章(都立大・院理)
16:30~18:30 ポスター
8月3日(火)
9:00~12:00 ポスター
10:00~11:00 奇数番号の説明
11:00~12:00 偶数番号の説明
13:20~16:00 シンポジウム
Symposium "Symmetry Breakers"
Organizers: Tadashi Uemura (Kyoto University)
& Fumio Matsuzaki (Tohoku University)
S-1 13:20 Tadao Usui (Kyoto University)
S-2 13:55 Shigeo Hayashi (National Institute of Genetics)
(14:30~14:45 break)
S-3 14:45 Fumio Matsuzaki (Tohoku University)
S-4 15:20 William Chia (National University of Singapore)
16:00~16:30 総会
18:00~20:00 懇親会
(会場:名古屋大学ユニバーサルクラブ、
会場での受付開始は17:00頃から)
8月4日(水)
9:00~11:00 ポスター
11:00~12:00 テクニカルセミナー
・RNAi
・GFPを用いた生きた胚のリアルタイム観察
・その他
(ポスター撤去は、12:45までに!)
13:00~15:00 討論会「ポストゲノム時代にむけて」
コーディネーター:伊藤 啓(基生研)
15:00 閉会
シンポジウム演題
8月3日(火)13:20~16:00
Symposium "Symmetry Breakers"
Organizers: Tadashi Uemura (Kyoto University)
& Fumio Matsuzaki (Tohoku University)
S-1 Flamingo: Multiple Roles in Planar Cell Polarity Formation
and Axon/Dendrite Patterning.
Tadao Usui*1, Yasuyuki Shima1, Yuko Shimada1, Shinji Hirano2,
Yuko Yamaguchi1, Robert W. Burgess3, Thomas L. Schwarz4,
Masatoshi Takeichi1, Tadashi Uemura1.
1:Graduate School of Biostudies, Kyoto univ.,
2:(Present address) Aichi Human Service Center,
3:Stanford univ. School of Medicine.
S-2 Cell specification and polarization in the tracheal development.
Shigeo Hayashi*, Tomoatsu Ikeya, Miho Tanaka-Matakastu,
Takahiro Chihara.
National Institute of Genetics and the Graduate School
for Advanced Studies.
S-3 Asymmetry in neural stem cells.
Fumio Matsuzaki.
Institute of Development, Aging and Cancer. Tohoku University.
S-4 Asymmetric cell divisions and the generation of neuronal diversity
William Chia.
Developmental Neurobiology Laboratory, Institute of Molecular
and Cell Biology, National University of Singapore.
講演演題
8月2日(月)13:05~16:30
<座長:井上 喜博(愛知がんセ研・生物)>
L-1 Notch シグナリングと器官アイデンティティーの決定
倉田 祥一朗
東北大・院薬
L-2 Dppモルフォゲンとパターン形成
多羽田哲也
東大・分生研
L-3 新規Ets蛋白EDLによるhomeogenetic induction の制御
山田琢磨1、岡部正隆1、広海健*1,2
1: 国立遺伝学研究所、2: 総合研究大学院大学・遺伝学専攻
<座長:中村 真(基生研・形態形成)>
L-4 ショウジョウバエを用いた細胞死実行因子の機能解析
三浦正幸
大阪大・院医・神経機能解剖
L-5 ショウジョウバエDNA複製関連遺伝子の転写制御ネットワーク
山口政光
愛知がんセ研・生物
L-6 オウトウショウジョウバエ亜群の雄が作る排卵刺激ペプチド
布山喜章
都立大・院理・生物科学
ポスター一覧
P-01 Dfrizzled-3, a new Drosophila Wnt receptor, acting as an attenuator of Wingless signaling.
佐藤 淳*1、小嶋 徹也1、程 久美子2、宮田 雄平2、西郷 薫1
1:東大・理・生化、2:日医大・薬理
P-02 細胞膜プロテオグリカン Dally の翅の発生における機能
藤瀬桃子*、泉 進、中藤博志
都立大・院理・生物
P-03 減数分裂突然変異体mei-yh92の細胞遺伝学的解析
平井和之*1、山本雅敏2
1:京都工繊大・院工芸科学・生物機能、
2:京都工繊大・ショウジョウバエ遺伝資源センター
P-04 D. melanogaster/D. simulans 雑種における不妊遺伝子のマッピング
澤村京一*、山本雅敏
京都工繊大・ショウジョウバエ遺伝資源センター
P-05 dally 遺伝子の転写及び転写後調節機構
津田 学*、泉 進、中藤博志
都立大・院理・生物
P-06 キイロショウジョウバエの卵成熟
初見真知子*1、澤正実2、渡辺泰弘3
1:島根大・生科・生物、2:愛教大・生物、3:島根大・理・生物
P-07 Notch シグナリングと器官アイデンティティーの決定
倉田 祥一朗*1,2、郷正博3、Spyros Artavanis-Tsakonas3,
Walter J. Gehring2
1: 東北大・院薬、2: Biozentrum, University of Basel, Switzerland、
3: HowardHughes Medical Institute, Yale University, USA
P-08 ショウジョウバエ変態中の中枢神経系における細胞死
辻村秀信*1、古賀裕美子1、木村賢一2
1:農工大・農・発生生物、2:北海道教育大
P-09 melanogaster species subgroupにおけるミトコンドリアDNAのA+T-rich領域の進化
辻野史*1、小瀬村暁子1、猪平佳代2、松浦悦子2
1: お茶大・人間文化・ライフサイエンス、2: お茶大・理・生物
P-10 細胞接着因子とStill life - Rac経路のシナプスにおける局在と機能
曽根雅紀*1, 2、鈴木えみ子2, 3、星野幹雄4、侯東梅5、黒見坦5、
深田正紀6, 7、黒田真也6、貝淵弘三6、鍋島陽一4、浜千尋1, 2
1 : 国立精神神経センター・遺伝子工学、2 : CREST、3 : 東大・医科研、
4 : 京大・医、5 : 群馬大・医、6 : 奈良先端大、7 : 広島大・医
P-11 ショウジョウバエ脳におけるキノコ体初期形成過程の分子遺伝学的解析
来栖光彦*、長尾智子、古久保-徳永 克男
筑波大学 生物科学
P-12 グリア・ニューロンを作りだす非対称性の分子機構
秋山(小田)康子*1、月田承一郎1,2、小田広樹2
1:京大・医・分子細胞情報、2:科技団・月田細胞軸プロジェクト
P-13 ショウジョウバエCaspaseファミリーの活性化機構
嘉糠洋陸*、澤本和延、岡野栄之、三浦正幸
阪大・医・神経機能解剖&CREST
P-14 ショウジョウバエcentral complexの蛹期における発生
田中公子*1,2、伊藤 啓1,3
1:基生研、2:CREST・JST、3:PRESTO・JST
P-15 ヒトp53をショウジョウバエ複眼原基で異所的に発現させるとS期への進入を阻害し、アポトーシスを誘導する
山口政光*1、広瀬富美子1、井上喜博1、白木岐奈1、林裕子1、西義美2、
松影昭夫1
1:愛知がんセ研・生物、2:愛知がんセ研・共通
P-16 神経-筋結合の標的認識過程における軸索成長円錐と筋細胞の動的相互作用
鈴木えみ子*1, Demian Rose2, Sarah Ritzenthaler2, 千葉晶2
1:東大・医科研・微細形態, CREST.JST,2:Dept. Cell & Struct.
Biol., Univ.Illinois
P-17 光受容細胞の分化におけるMSIおよびSINAの機能
廣田ゆき*1、岡部正隆2、来栖光彦3、澤本和延1、岡野栄之1
1:阪大・医・神経機能解剖学、CREST、2:遺伝研・発生遺伝研究部門、
3:筑波大・バイオシステム
P-18 ショウジョウバエを用いた無脊椎動物Otx 遺伝子の機能解析
安達在嗣*1、長尾智子1、西駕秀俊2、阿形清和3、梅園良彦3、Hans Bode4、
古久保ー徳永克男1
1:筑波大・生物、2:都立大・生物、3:姫工大・生命、
4:Dept. Dev. Cell Biol.,Univ. Ca lifornia
P-19 末梢神経系におけるNotchシグナルを介したgcm遺伝子の発現調節
梅園良彦*1、細谷俊彦2,3、堀田凱樹1,3
1:科技団・CREST、2:科技団・さきがけ研究21、3:国立遺伝研・発生遺伝
P-20 アクチン細胞骨格系を制御する新規MAPキナーゼフォスファターゼ
丹羽隆介*1、Bruce Hay2、竹市雅俊1、上村匡1
1:京大院・生命科学、2:カリフォルニア工科大
P-21 Notchシグナリングによるプロニューラル蛋白アキートの機能抑制
中尾啓子*、Fisher, A., Caudy, M.
コーネル大学医学部
P-22 母性効果致死遺伝子tibi はArp2/3 complex 21kd subunitをコードしている
松林宏*1、山本雅敏2
1:京都工繊大学・院、2:京都工繊大・ショウジョウバエ遺伝資源センター
P-23 One-hybrid法によってクローン化されたショウジョウバエPCNA遺伝子転写制御因子Grainyhead(GRH)
林 裕子1*、山岸 正裕1、西本 義男1、田口 修2、松影 昭夫1、
山口 政光1
1: 愛知がんセ・研・生物 2: 愛知がんセ・研・二病
P-24 キイロショウジョウバエ処女雌の性的受容性を制御するchaste 遺伝子の解析
従二 直人*1、山元 大輔1,2
1:科技団・山元行動進化プロジェクト、2:早大 ・人間科学部
P-25 Identification and characterization of Drosophila homolog of Rho-kinase.
Tomoaki Mizuno1*, Mutsuki Amano2, Kozo Kaibuchi2 and
Yasuyoshi Nishida1.
1: Grad. Sch. of Sci., Nagoya Univ., 2: Nara Inst. of Sci. and
Technol.
P-26 カブトムシの雄特異的角形成の分子マーカーの検索
新美輝幸*、大島宏之、三輪雅代、山下興亜
名大院・生命農
P-27 Calreticulin 遺伝子突然変異体における可塑的行動の異常
冨田純也*1、国吉久人2、従二直人2、蒲生寿美子1、山元大輔2,3
1:大府大・総合科学研究科、2:科学技術振興事業団・山元行動進化プロジェクト、
3:早大・人間科学部
P-28 ショウジョウバエ個体を用いたヒトアミロイド前駆体蛋白質(APP)の過剰発現表現型の解析
八木克将*1,2、富田進1、中村真3、桐野豊1、鈴木利治1
1:東大・薬・神経生物物理、2:生研機構、3:基生研
P-29 キイロショウジョウバエの交尾時間が不規則になる変異体 fickle の解析
馬場浩太郎*1、竹下綾2,3、従二直人2、山元大輔2,4
1:東大・理・物理、2:科学技術振興事業団・山元プロジェクト、
3:現:三菱化学生命研、4:早稲田大学・人間科学部
P-30 DERシグナル因子ebi はNotchシグナルと相互作用を示す
津田 玲生*、S. Lawrence Zipursky
HHMI, カリフォルニア大学ロサンゼルス校
P-31 WGAトランスジーンを用いたショウジョウバエ神経筋接合部におけるシナプス形成機構の解析
森本(谷藤)高子*1、徳本貴久1、田渕克彦2、岡野栄之2、吉原良浩3、
能瀬聡直1
1:東京大・院理・物理、2:大阪大・医・神経機能解剖、
3:理研・脳研・シナプス分子機構
P-32 ショウジョウバエ成虫原基における異所性感覚器官形成と位置情報の関係
丹羽尚*、岡部正隆、広海健
国立遺伝研・発生遺伝
P-33 異所発現トラップ法を用いた神経結合特異性に関わる新規遺伝子の探索
梅宮猛1,2、高須悦子1、田中宏昌1、竹市雅俊2、相垣敏郎3、能瀬聡直*1
1:東大・院理・物理、2:京大・院理・生物物理、3:都立大・生物
P-34 グリア細胞は複数の分化経路で分化する
滝沢 一永*、堀田 凱樹
遺伝研、発生遺伝 科学技術振興事業団
P-35 交尾における生殖器の連結および解除に異常を示すキイロショウジョウバエの変異体、lingerer の解析
国吉久人*1、馬場浩太郎2、近藤俊三3、山元大輔1,4
1:科技振、山元プロジェクト、2:東大理学部、3:三菱化学生命研、
4:早大人間科学部
P-36 複眼光受容細胞のニューロン分化におけるネガティブレギュレーターの役割
岩波 将輝*1.2、広海 健1
1:国立遺伝学研究所 発生遺伝研究部門 2:慈恵医大 微生物 第1
P-37 ショウジョウバエ翅・脚原基形成におけるEGFRシグナルの役割
久保田 一政*1,2、後藤 聡2,3、江藤 一洋1、林 茂生2,3
1:東医歯大・歯・発生、2:国立遺伝研、3総研大
P-38 オナジショウジョウバエのpremating isolationの遺伝学的研究
上野山 登久
神戸学院女子短大
P-39 FTZ-F1とFTZの相互作用機構
鈴木 大河1、梅園 和彦1、川崎 陽久2、上田 均*2
1:京大・ウイルス研・情報高分子化学、2: 遺伝研・形質遺伝
P-40 小笠原諸島のオナジショウジョウバエはヤエヤマアオキ果実で繁殖可能か?
梁 百霊*、布山喜章
都立大・院理・生物科学
P-41 アドヘレンスジャンクションのネットワーク形成:カドヘリンとカテニンの協調的な作用
小田広樹*1、月田承一郎1, 2
1: ERATO・月田細胞軸プロジェクト、2: 京都大・医
P-42 染色体分配を制御する新しい遺伝子(orbitとmeteor)の同定とその機能の解析
井上喜博*1、山口政光1、西本義男1、D. M. Glover2、松影昭夫1
1:愛知がんセ研・生物、2:Dept. of Genetics, Cambridge Univ. England
P-43 神経系に特異的に発現するC2HCタイプZnフィンガー転写因子の構造と機能
大迫俊二*、高松芳樹
東京都神経研・細胞生物
P-44 ハエの小容器内での歩行活動は2つの相からなる
小松 明
東京女子医大・医・第一生理
P-45 hedgehog 及びdpp のモルフォゲン情報を統合する新規パターン形成遺伝子のクローニング、及び機能解析
船越陽子*、南真樹、多羽田哲也
東大・分生研
P-46 ショウジョウバエ気管系における Wg シグナリングの二つの機能
千原崇裕*、林茂生
遺伝研・無脊椎、総研大・生命科学
P-47 キノコ体を中心としたCaM キナーゼII - GAL 4の発現部位の解析
高松芳樹*1、中越英樹2、西田育巧3、山内卓4、大迫俊二1
1: 都神経研・細胞生物、2: 東工大・フロンティア(現岡山大・理・生物)、
3: 名大・理・生物、4: 徳島大・薬・生化
P-48 Notch 情報伝達系構成因子の新規候補遺伝子である aya の解析
鈴木 聡*1、伊藤美紀子 2、岸 憲幸 1、松野康子 3、
Spyros Artavanis - Tsakonas、3、岡野栄之1 ,2 、松野健治 2→4
1 : 大阪大学大学院医学系研究科・神経機能解剖、
2 : 科学技術振興事業団/CREST/JST、3 : Cell Biol.、HHMI ,
Sch. of Med. Yale , USA、 4 : 東京理科大学基礎工学部生物工学科
P-49 Notchシグナルが果たす気管での役割
池谷智淳*1 林茂生1,2
1:遺伝学研究所、2:総合大学院大学
P-50 キイロショウジョウバエ雌の交尾のコストに及ぼす卵の有無の影響
上山盛夫*、布山喜章
都立大・院理・生物科学
P-51 ショウジョウバエSTATはD-raf プロトオンコジーンの転写制御因子である
権恩貞*1, 2, 3, 朴賢淑2, 金英信2, 西田育巧3, 松影昭夫1, 劉美愛2 ,
山口政光1
1愛知がんセンター研・生物、2釜山大・理・分子生物、3名大・院理・生命理学
P-52 DRab2は新生ロドプシンの輸送と網膜の色素顆粒形成の両方に必要である
佐藤明子*・河村悟・尾崎浩一
阪大・院理・生物
P-53 Gal4エンハンサートラップ系統の作出と翅成虫原基での発現スクリーン
山本美智子*1,山下敦士1,橋本直子1,高橋美和1,2,上田龍1
1:三菱生命研・神経発生遺伝,2:お茶大・人間文化
P-54 ショウジョウバエ消化管における左右非対称性
林 知美*、村上柳太郎
山口大・理・自然情報
P-55 初期胚での高い発現と染色体恒常性;ショウジョウバエRECQホモログ
川崎勝己*1,2, 鄭相民1, NGUYEN, Quang D. 1, 柴田武彦1,2,
1:理研 遺伝生化学、2: CREST, JST
P-56 ナイドジェン:ショウジョウバエ発生における細胞外マトリクスの解析
熊谷知乃1、Liselotte I. Fessler1, Stefan Baumgartner2,
John H. Fessler1
1 :UCLA, Mol. Biol. Inst. & MCD-Biol. Dept., USA,
2: Lund Univ., Dept. Cell & Mol. Biol., Sweden
P-57 シナプス小胞はrab5非依存的に再形成される
志水英之*、佐藤明子、河村悟、尾崎浩一
阪大・院理・生物
P-58 ショウジョウバエ変態過程におけるプログラム細胞死:細胞死抑制因子p35の強制発現による細胞死の抑制
児玉明聡*1、谷村禎一2、木村賢一1
1:北教大・岩見沢・生物、2:九大・理・生物
P-59 アナナスショウジョウバエ類の性的隔離における求愛歌の役割
山田博万*、都丸雅敏、小熊譲
筑波大・生物科学
P-60 ショウジョウバエRab蛋白質の一種、DRabRP1、の細胞内局在と特性
藤川和世*・河村悟・尾崎浩一
大阪大・院理・生物
P-61 キイロショウジョウバエとセイシェルジョウバエとの間の性的隔離と雌による配偶者選択の種内変異について
都丸雅敏*,小熊 讓
筑波大・生物科学
P-62 Dsrc42は上皮細胞嚢形成においてDJNKを調節する
館野実*、安達卓、西田育巧
名大・院理・生命
P-63 キイロショウジョウバエ自然集団における殺虫剤抵抗性の遺伝的変異と季節変動
三代隆洋*1、赤井住郎2、小熊讓3
1:筑波大・院・生物、2:山梨学院短大、3:筑波大・生物
P-64 ネトリン受容体フラッツルドはネトリンの局在パターンを変化させ、細胞非自律的に軸索ガイダンスを行う
平本 正輝*、堀田 凱樹
遺伝研・発生遺伝
P-65 Drosophila mab-21の機能解析
白木岐奈*1,2、高橋直樹1、林茂生2
1:奈良先端大・バイオ・動物代謝、2:遺伝研・無脊椎
P-66 強制発現系を用いたショウジョウバエ寿命変異体の探索
成 耆鉉*1、相垣敏郎1,2
1:都立大・院・理、2:科技団さきがけ)
P-67 オーストラリア東海岸におけるキイロショウジョウバエのゲノム内のP因子の変遷
小倉啓司*1、Ronny C. Woodruff 2、伊藤雅信 3、Ian A. Boussy 4
1: 工繊大・ショウジョウバエ遺伝資源センター、2: Dept. of Biol. Sci.,
Bowling Green State Univ. 、3: 工繊大・繊維・応用生物、
4: Dept. of Bio., Loyola Univ. of Chicago
P-68 red 遺伝子の転写産物の同定
大迫隆史1、相垣敏郎1,2、布山喜章1
1:都立大・理・生物、2:科技団・さきがけ
P-69 核マトリックス構成分子Plexusの翅脈パターン形成での役割
亦勝和*1,2、田所竜介1、蒲生寿美子2、林茂生1
1:国立遺伝研、2:大阪府立大学・総科、
P-70 減数分裂突然変異体mei(3)1223[m144]による染色体特異的対合機構の解析
山本雅敏*1、平井和之2
1:京都工繊大・ショウジョウバエ遺伝資源センター、
2:京都工繊大・院工芸科学・生物機能
P-71 肢原基で発現するgal4エンハンサートラップ系統のスクリーニング
後藤聡*1,2、谷口美佐子1、林茂生1,2
1: 国立遺伝研・系統セ・無脊椎、2: 総研大
P-72 The GFP viewer: A low-cost DIY portable device for the observation of Green Balancers and transgenics S65T GFP expression.
ペール JB*1、相垣敏郎1,2
1: 東京都立大学・理学・生物、2: JST
P-73 HedgehogによるDppシグナリングの負の制御
谷本拓*、多羽田哲也
東大・分生研
P-74 神経回路の形成を制御するdrio遺伝子の解析
粟崎健1、酒井良子、斉藤麻衣、曽根雅紀、浜千尋*
国立精神神経センター神経研究所遺伝子工学研究部、
1:現、国立基礎生物学研究所
P-75 幼虫の視神経細胞の分化に働く新しいhedgehogシグナル経路
鈴木崇之、西郷薫
東大・理・生化
P-76 Dppシグナル伝達に関与する新規因子の遺伝学的スクリーニング
中村 真*、西田 弥生、友安 慶典、上野 直人
基礎生物学研究所・形態形成研究部門
P-77 ショウジョウバエの巨大筋肉蛋白質Kettinの構造および遺伝的解析
羽毛田 聡子*、西郷 薫
東大・理・生化
P-78 間接飛翔筋由来cDNAライブラリーの作成
最上 要*
東大・院理・物理
P-79 wing disc形成に異常を示す新規変異体 unbalanced flight の解析
友安 慶典*1,2、上野 直人1,2、中村 真1
1:基生研・形態形成、2:総研大
P-80 FTZ-F1 変異体による fushi tarazu 遺伝子転写調節の解析
川崎陽久*1,2、上田均1、広瀬進1
1:遺伝研・形質、2:岩手大・連合農学
P-81 キイロショウジョウバエDNA polymeraseδ及びε欠損突然変異体の単離の試み
吉田英樹*1、2、井上喜博1、山口政光1、広瀬富美子1、大重真彦2、
坂口謙吾2、松影昭夫1
1:愛知県がんセ研・生物、2:東理大・理工・応用生物
P-82 細胞の神経誘導能獲得に関与するショウジョウバエの遺伝子 edl
山田 琢磨*1、岡部 正隆1、三田和英2、広海 健1
1:国立遺伝研・発生遺伝 2:放医研・ゲノム
P-83 ショウジョウバエ新規rasファミリー遺伝子Rap2-likeの分子生物学的解析
宮田直政*1、大迫隆史1、相垣敏郎1,2
1:都立大・院理・生物科学、2:科技団・さきがけ
P-84 シナプスの可塑性、学習に寄与するショウジョウバエlinotte遺伝子
齊藤 実*1、Tim Tully2
1:東京都神経研・病態神経生理、2:コールドスプリングハーバー研究所
P-85 cAMPによるシナプス可塑性の遺伝解析
吉原基二郎*、城所良明
群馬大学医学部行動医学研究施設
P-86 翅縁形成に関与する遺伝子 hiiragi の機能解析
長曽秀幸*1、村田武英1、岡野栄之2、横山和尚1
1:理研・筑波セ、2:阪大・医・神経機能解剖学、科技団(CREST)
P-87 DREF結合因子として分離したショウジョウバエMLFホモログの機能解析
大野勝人*1,2、高橋康彦3、広瀬富美子1、井上喜博1、田口修4、西田育巧2、
松影昭夫1、山口政光1
1:愛知がんセ・研・生物、4:二病、2:名大・院理・生命、3:ハーバード大
P-88 ショウジョウバエのヘモサイチン遺伝子
後藤彰*1、熊谷剛1、森仁志1、森肇2、北川泰雄1,3
1:名大院・生命農・生化学制御、2:京都工繊大・繊維・応用生物、
3:名大・生物分子応答研究セ
P-89 全身麻酔薬への感受性に関係する遺伝子群の解析
蒲生寿美子1*, 田中良晴1, 亦勝和1、石井秀紀1, 冨田純也1, 佐子山豈彦2,
1: 大阪府大・総合, 2: 阪大・医・遺伝
P-90 ショウジョウバエの新規プロテアーゼインヒビターファミリーの遺伝子解析
横山裕昭*1、新美友章2、Konrad Beck2、北川泰雄1、2
1:名大院生命農学・生化学制御、2:名大・生物分子応答研究セ
P-91 Argosシグナル伝達経路に関与する新規な遺伝子の同定と機能解析
田口明子*1、澤本和延1, 2、宮尾幸代1, 2、岡野栄之1, 2
1: 阪大・医・神経機能解剖学、2:CREST
P-92 Transcriptional Regulation of Drosophila TATA-Box Binding Protein (TBP) Gene by DRE/DREF system and Homeodomain Protein Zen.
Tae-Yeong Choi1*, Kwang-Hee Baek2, Jae-Seong Yoon2,
Kyu-Hyung Han3, Akio Matsukage4 and Mi-Ae Yoo1 .
1:Dept. of Mol. Biol., Pusan National Univ., Pusan, Korea;
2:Dept. and Inst. of Genet. Engineer., Kyung Hee Univ., Suwon, Korea;
3:Dept. of Genet. Engineer., Hallym Univ., Chunchon, Korea;
4:Lab. of Cell Biol., Aichi Cancer Center Res. Inst., Nagoya, Japan .
メンデルの足跡を訪ねて
伊藤 啓 (基礎生物学研究所)
(写真をクリックすると大きな絵が見られます。)
土俗宗教に対する近代宗教の定義は、教祖、教義、布教組織の3つが存在することだという。だが言い出しっぺが居り、ドグマがあり、それを広める組織があれば宗教と呼べるのであれば、ほとんどの学問は実は、宗教の定義にあてはまってしまう。ニュートンを祖とする古典力学教、ダーウィンを祖とする進化論教など、大学という布教組織において、教授という司祭によって、今も活発に布教が続けられている。教祖の言うことを盲信するのでなく批判的に発展させていくのが宗教と学問の違いだという意見もあるが、仏教だってキリスト教だって、いまの内容が教祖の最初の教えとはずいぶん違ったものに発展している点では、進化論などとさほど変わりはない。
さて、これらの中にあって遺伝学という宗教は、その教祖が不遇のうちに注目されぬまま一生を終え、死後しばらくたってから劇的なカムバックを遂げたあたり、キリスト教にも似たところがあって、実にドラマチックな要素を備えている。ショウジョウバエの研究者としてこの宗教の布教組織の末端に連なっている人間にとって、教祖の生地、活動の地を訪ねて足跡を偲ぶことは、イスラム教徒のメッカ巡礼同様、一生に一度はやらねばならぬ聖なる義務と言っても過言ではなかろう。このような次第で、ドイツでポストドク生活を始めて1年ほどたった1992年の夏、車を手に入れてから最初に迎えたバカンスシーズンは、我らが教祖メンデルゆかりの聖地を巡礼してみることに決めたのであった。
(というのは全くのこじつけで、本当はチェコにいる知人がたまたまメンデルの修道院がある町に住んでいただけの理由なのだが。)
ドイツ人の休暇は概して長く、研究室のメンバーも教授から技官までみな3週間~1ケ月の休みを取る。新参の日本人ポストドクとしては、ちょっと遠慮して2週間だけ休むことにした。これだけあれば、メンデルの修道院があるブルノだけでなく、生まれた村であるハインツェンドルフ、高校に通ったオロモウツ、そして大学で学んだウィーンをすべて回ることができる。私が住んでいたマインツという町は、ドイツの中でも西のはしに近い。メンデルの故郷であるハインツェンドルフは、隣国チェコの東のはじである。だが直線距離ではせいぜい650キロ。細長い日本に比べると、ヨーロッパは狭い。
チェコに行くには、まずアウトバーンを飛ばして、ドイツを東のはじまで横断する必要がある。アウトバーンは承知のとおり速度無制限であるが、私が茶目っけを出して買い込んだ車は、1948年以来変わらぬデザインで40年間製造が続けられていたシトロエン2CVというクラシックカーに近い代物で、28馬力を誇る二気筒エンジンでは、どうアクセルを踏んでも110キロしか出ない。200キロで飛ばす方々の邪魔にならぬよう、はしの車線を軽飛行機のような爆音を立てつつそろそろと走る。国境手前の町まで来たところで、1日目は終った。
1989年のビロード革命で共産圏を脱してから間もない当時のチェコでは、入国にはビザが要った。飛行機や列車で入国する場合、あらかじめ他国の首都にある領事館で長い行列をして、ときには交付まで数日間待たされて、ビザを取得しなくてはならない。ところが自動車の場合は、幾つかの大きな幹線道路を選んで通れば、国境の検問所のすぐ脇の事務所でその場でビザを取ることができる。手数料5000円ほどを払えばすぐにスタンプを押して入場ゲートを通してくれるあたり、何となくディズニーランドの入口で「パスポート」券を買うときと同じ気分である。
国境を越えると景色はがらっと変わる。黒くごついドイツの森に比べ、チェコの森は(フランスもそうだが)明るく柔らかい感じがする。何でもこれは、ドイツでは植林してから40年は木を切らないのに、他の国では20~30年で切ってしまうかららしい。地形というものは変えようがなくても、景色というものは人の営みでずいぶん変わるものである。林業という意味ではドイツの方が優れているのかも知れないが、見た目という点ではチェコの方が目に優しい。
メンデルはチェコの東部、モラビア地方の出身である。ドイツ国境からはさらに、チェコをほぼ横断する必要がある。ところが途中には、宇宙から帰還したガガーリンもリハビリに来たという温泉の名所であるカルロビバリ(昔はカールスバートと呼ばれ、ゲーテやらメッテルニヒやらも保養に来ていた)、地球で一番おいしいビールの産地であるピルゼン(日本製ビールの大半を占めるピルスナータイプは、この町のビールの製法に従ったビールの総称である)、そしてヨーロッパでも最も美しい街の一つであるプラハなどの「障害物」があり、旅はなかなか先へ進まない。
寄り道を終えて、やっとモラビア地方に入る。まず最初の目的は、メンデルが生まれた村を見つけることである。メンデルがいた修道院はガイドブックにも載っている観光名所であるが、生まれた村は全く有名でない。唯一の手がかりは、岩波文庫「雑種植物の研究」(旧版)の付録についている「メンデル小傳」という文章の中に書かれた、「モラヴィアの北の端、今日では獨逸、波蘭及びチェコ・スロワキアが境を接してゐるところに・・・小都會のオドラウ Odrau から北へ一時間歩程(4、5キロ)、舊い都會のノイチッチャイン Neutitschein から三時間歩程のところにある、ハインツェンドルフ Heinzendorf が、グレゴール・ヨーハン・メンデルの故郷である。」という一節だけである。生家が残っているのかも分からない。だが戦争で完全に破壊された家でも昔どおりに建て直して「キュリー夫人の生家」とか「バッハの生家」とかいうプレートを貼り付けたがるヨーロッパのことだから(しかもバッハの家なんて、本当は生家の隣の家でしかない)、きっと何かしら残っているだろうということは期待できる。
問題は、ハインツェンドルフという地名が地図のどこにも見当たらないことである。メンデルの時代、このあたりはオーストリア・ハンガリー帝国の一部で、ドイツ語が公用語であり、地名もドイツ語ふうのものが用いられていた。しかし第一次大戦でオーストリアが負けてチェコが独立してからは、たとえば「カールの風呂」を意味するカールスバートが同じ意味のチェコ語であるカルロビバリに直されたように、多くの地名が相当するチェコ語に直されたのである。ハインツェンドルフも明らかに「ハインツの村」というドイツ語だから、チェコ語に直されてしまったに違いないのだが、残念ながらどういうチェコ語になるのかが分からない。
ノイチッチャインも、やはりどこにあるのだか分からないのだが、さいわいもう一つの地名「オドラウ」は、Odrau が Odry に変わっただけで国のはずれ、ポーランド国境に近いあたりに見つかった。町の名前だけでなく、あたり一帯の郡の名前にもなっているらしい。そこでとりあえずこの町まで行き、そこから北4キロあたりでそれらしい村を探すことにする。
ガソリンスタンドと数十メートルの商店街からなる「小都會」オドラウの町を過ぎ、北のあたりを走り回る。道ばたの案内地図を見たり、道ゆく人を呼びとめて尋ねてみたりしても、ハインツェンドルフらしき地名には行きあたらない。チェコはオーストリア時代が長く、第二次大戦ではドイツに併合されていたうえ、40年間の共産圏時代も、押しつけられた同盟の盟主であるソ連のロシア語は嫌われ、ドイツ語が依然もっともポピュラーな外国語であった。だからハインツェンドルフがどういう地名に変わったかも、近所まで来ればすぐわかるだろうと思ったのが間違いだった。プラハのような観光都市と違い、さすがにこんな田舎まで来ると、ドイツ語が分かる人も多くはない。事前にもっと詳しく調べてくるべきだったと悔やまれる。
 2時間ほど走り回り、本当の村の場所は分からなかったけれど、まあこのような景色を見ながらメンデルは育ったのだろうということだけで満足して帰ろうかと思いだしたころ、小さな食堂を見つけた。昼下がりで、近所の人らしいのが15人ほど新聞を読んだり世間話をしたりしている。片端から聞いてみれば誰か一人くらいは知っているかもしれないと考え、店の中に入ってみた。たいへん運の良いことに、客の中にドイツ語が分かり、しかもタクシー運転手をしているという願ってもない人がいた。ああ、それなら多分ヒンツィーチェ Hyncice だ。オドラウのちょっと南だよ。と差し出した地図に印をつけてくれた。
2時間ほど走り回り、本当の村の場所は分からなかったけれど、まあこのような景色を見ながらメンデルは育ったのだろうということだけで満足して帰ろうかと思いだしたころ、小さな食堂を見つけた。昼下がりで、近所の人らしいのが15人ほど新聞を読んだり世間話をしたりしている。片端から聞いてみれば誰か一人くらいは知っているかもしれないと考え、店の中に入ってみた。たいへん運の良いことに、客の中にドイツ語が分かり、しかもタクシー運転手をしているという願ってもない人がいた。ああ、それなら多分ヒンツィーチェ Hyncice だ。オドラウのちょっと南だよ。と差し出した地図に印をつけてくれた。
Heinzendorf と Hyncice では似ても似つかぬようにみえるが、言われてみればハインツとヒンツィーは何となく似ているし、○○チェという地名はチェコには多く、ドイツ語の○○ドルフ(村)にあたると言われればたしかに納得できる。この地名なら地図にもちゃんと出ている。オドラウの「北」という教科書(岩波文庫)の記述を鵜呑みにせず、広い視野で東西南北くまなく探していれば良かったのだ。印刷された記載を無意識のうちに無批判に受け取ってしまった点、科学者としての修行がまだまだ足りないと反省した。
 南下してオドラウに戻り、もう2キロほど街道を南下してから、それて脇道に入る。さらに2キロほど行くと、「オドラウ郡ヒンツィーチェ」の看板が道端に見つかった。嬉しさのあまり思わず車を停めて、しばし感慨に浸る。ヒンツィーチェの村は、ゆるやかな起伏の畑地のなかに、ちょっと盛り上がった小さな丘の上にあった(写真1)。一本道がいったん坂を下り、村に続く丘のふもとに達したあたりの、上り坂になる少し手前の村の入口に、くすんだ茶色が普通のこのあたりの民家と異なり、やけに立派な白塗りの小屋があった(写真2)。屋根には豪勢な破風がつけられ、ドイツ語の花文字が書かれた石版が飾られている(写真3)。読んで見ると:
南下してオドラウに戻り、もう2キロほど街道を南下してから、それて脇道に入る。さらに2キロほど行くと、「オドラウ郡ヒンツィーチェ」の看板が道端に見つかった。嬉しさのあまり思わず車を停めて、しばし感慨に浸る。ヒンツィーチェの村は、ゆるやかな起伏の畑地のなかに、ちょっと盛り上がった小さな丘の上にあった(写真1)。一本道がいったん坂を下り、村に続く丘のふもとに達したあたりの、上り坂になる少し手前の村の入口に、くすんだ茶色が普通のこのあたりの民家と異なり、やけに立派な白塗りの小屋があった(写真2)。屋根には豪勢な破風がつけられ、ドイツ語の花文字が書かれた石版が飾られている(写真3)。読んで見ると:
傑出した自然科学者かつ植物学の権威である
グレゴール・ヨーハン・メンデル司教
生地の名誉市民にして消防団の創始者
を記念して
生:1822年7月22日 ハインツェンドルフ58番地
没:1884年1月6日 ブリュン
1902年建立
とある。
 日本でも村の入口には、よく「○○消防団」と書かれた小さな倉庫のような建物が建っている。村との位置関係といい建物の大きさといい、ちょうど同じである。例の岩波文庫の「メンデル小傳」には、「彼がいよいよ有名になった時に、新しい消防署の建立が、彼を記念する機會として用ひられて、記念碑をその建物に掲げて表彰した」とある。消防団にしては屋根だけがやけに立派なのが、面白い。開いていた扉から中を覗くと、荒れ果てた物置きになっていてほとんど使われていない様子であった。
日本でも村の入口には、よく「○○消防団」と書かれた小さな倉庫のような建物が建っている。村との位置関係といい建物の大きさといい、ちょうど同じである。例の岩波文庫の「メンデル小傳」には、「彼がいよいよ有名になった時に、新しい消防署の建立が、彼を記念する機會として用ひられて、記念碑をその建物に掲げて表彰した」とある。消防団にしては屋根だけがやけに立派なのが、面白い。開いていた扉から中を覗くと、荒れ果てた物置きになっていてほとんど使われていない様子であった。
過去の偉大な科学者100人の評伝を纏めた「The Scientific 100」(John Simmons 著 Citadel Press)という本によると、メンデルが遺伝の原理の発見者として再評価されるに至った経緯は、それほどナイーブなものでもなかったらしい。科学の歴史の中には、再発見して大々的に広めた人間の方が最初の発見者よりも後世有名になっている例は少なくない。だが1900年には、3人の科学者が独立に遺伝の原理に関する研究を発表しようと競争していた。現代と同じで、当然そこにはプライオリティに関する争いが生じる。すでに死んだ人間であるメンデルを遺伝の原理の発見者として担ぎ出し、ことさらに称賛することには、そういう争いを回避しようという思惑も多少はあったのだろう。
メンデル再発見に対するドイツ圏の反応には、1900年という時期も大きく影響していたに違いない。1900年といえば、鉄血宰相ビスマルク率いるプロシャが1891年に普仏戦争でフランスを破り、統一ドイツを樹立してからわずか十年。基礎科学の分野でのドイツ圏の飛躍的な活躍が最高潮のころである。こういう中で、生物学の流れを根底から変えるような大発見が、イギリスやフランスではなくドイツ圏でなされていたことが明らかになったというのは、決して悪い気分がすることではない。そんなわけで、やけに立派な記念碑のような屋根を持つ消防団詰所というミスマッチな建物が残されることになったのではなかろうか。建物の周りを何周もうろうろしながら、そんな事を考えた。
 丘の上の村へ続くカーブした坂をのぼり切ると、まっすぐな道の両側に数十軒ほどの家が並ぶヒンツィーチェの村に出る。家並みの真ん中へんには道の片側に教会が立っていて、そのへんが村の中心らしい。メンデルの家はこの村の58番地、58番地と言えば坂をのぼり切ってすぐ、家並みの一番はずれのあたりである。その位置には茶色い三角屋根の家が建っている。道路に面した壁の中程には、小さなプレートが埋め込んである。こんどはチェコ語なので何と書いてあるのかよく分からないが、Mendel という文字は判読できる(写真4)。これが生家だと信じてよさそうだ。180年前の家がそのまま残っているのかどうかは確かめられなかったが、チェコはほとんど戦災にあっていないから、その確率は高い。
丘の上の村へ続くカーブした坂をのぼり切ると、まっすぐな道の両側に数十軒ほどの家が並ぶヒンツィーチェの村に出る。家並みの真ん中へんには道の片側に教会が立っていて、そのへんが村の中心らしい。メンデルの家はこの村の58番地、58番地と言えば坂をのぼり切ってすぐ、家並みの一番はずれのあたりである。その位置には茶色い三角屋根の家が建っている。道路に面した壁の中程には、小さなプレートが埋め込んである。こんどはチェコ語なので何と書いてあるのかよく分からないが、Mendel という文字は判読できる(写真4)。これが生家だと信じてよさそうだ。180年前の家がそのまま残っているのかどうかは確かめられなかったが、チェコはほとんど戦災にあっていないから、その確率は高い。
メンデルの家は、左半分が納屋になっていてそちら側に入り口があった。村の家並みの中でもまあまあの大きさの家である(写真5)。裕福ではないがまあ貧しいほうでもない農家だったのだろう。いまどういう人が住んでいるのか分からないが、閉じている門をノックして訪ねてみるだけの度胸はなく、しばし家の前でたたずみ、ヨーハン・メンデル少年が行き来したであろう村の一本道をうろうろしてみたのち、ヒンツィーチェの村をあとにした。
 成績が良かったメンデルは、学校の先生の薦めもあり、ギムナジウムを経て大きな町の高校(日本の旧制高校に当たる)へ進学することができた。北部モラビア随一の町は、当時はオロミュッツ、現在はオロモウツと呼ばれる、このあたりの司教座聖堂もある町である。(ちなみにベートーベンの「戴冠ミサ曲」は、彼の友人がウィーンからオロミュッツの司教になって赴任した際に書いた曲である。)メンデルに従い、オロモウツへ向かう。ヒンチーチェからは約50キロ、当時はまる1日以上の旅程だっただろう。当然メンデルは寄宿生活を送っていたはずだ。父の病気で仕送りも滞りがちになり、しかも自身も3回も病気を患って、必ずしも明るく楽しい高校生活ではなかったようである。
成績が良かったメンデルは、学校の先生の薦めもあり、ギムナジウムを経て大きな町の高校(日本の旧制高校に当たる)へ進学することができた。北部モラビア随一の町は、当時はオロミュッツ、現在はオロモウツと呼ばれる、このあたりの司教座聖堂もある町である。(ちなみにベートーベンの「戴冠ミサ曲」は、彼の友人がウィーンからオロミュッツの司教になって赴任した際に書いた曲である。)メンデルに従い、オロモウツへ向かう。ヒンチーチェからは約50キロ、当時はまる1日以上の旅程だっただろう。当然メンデルは寄宿生活を送っていたはずだ。父の病気で仕送りも滞りがちになり、しかも自身も3回も病気を患って、必ずしも明るく楽しい高校生活ではなかったようである。
オロモウツは立派な大聖堂を中心に豪勢な石造りの建物が数多く立ち並び、それらが排気ガスのためか酸性雨のためかみなたいそう黒ずんでいて、恐ろしく古めかしい印象をかもしだしていた。さすがにかなり大きな町で、150年前はともかく今では無数の高校がありそうだ。メンデルの通った学校を探すのはあきらめた。
1843年、高校を卒業した21歳のメンデルは修道士への道を選び、モラビア地方の首都であり、現在でもチェコ第二の都市(といってもナンバーツーの常で、プラハよりははるかに小さいが)であるブリュン(現在はチェコ語でブルノ)の修道院に入った。修道院は単に隠棲する修行僧が住む場所というのではなく、地域の初等中等教育を統括し、農業の技術指導も行うなど、大学に准ずる学術文化の中心でもあった。とくに当時のブルノの修道院長であったナップ司教は、優秀な研究者肌の人材を集めていたらしい。大学に進むだけの資力はなかったメンデルにとって、修道院に入って学問を続けることができるのは、故郷に帰って農業をするよりは望ましい選択だったのだろう。彼はここで神学の講義を受けるとともに、近在のギムナジウムで数学やラテン語ギリシャ語を教えた。5年後には一人前の修行僧としての受戒をうけ、グレゴールというホーリー・ネ-ムをもらう。
 ブルノはオロミュッツより70キロほど南の、モラビア地方の南端にある町である。チェコの中といっても、プラハまでは200キロもあるが、真南のウィーンまでは110キロしかない。オーストリア・ハンガリー帝国時代には今よりもいっそう、帝国の首都との連絡は密だったに違いない。メンデルは理科の正式教員の免許を取るため、1850年にウィーンに赴いて教員資格試験を受けるが落ちてしまう。だが試験委員長には良い印象を持たれたらしく、ウィーン大学への留学を勧められ、修道院長の理解もあって修道院派遣の学生として2年間の留学を認められた。メンデルは主に物理学を学び、指導教官は(救急車のサイレン音の研究で名高い)ドップラーであった。1853年に留学を終えて修道院に戻ったメンデルは、開設されたばかりの工業学校で実験物理を教えはじめ、その仕事は彼が修道院長になるまでずっと続けられた。帰還3年後の1856年から始められたエンドウ豆を中心とする遺伝の実験が、当時の生物学の仕事としては異様なほど緻密な実験計画と論理的考察に基づいて進められているのは、彼の物理のバックグラウンドが間違いなく影響しているのであろう。1865年に発表された彼の論文が、別刷りを世界中の著名な生物研究者に送りつけるという彼の努力にも関わらず当時ほとんど理解されなかったのは、ある意味では無理もない。物理学者の間には、化学や地学や生物の連中が膨大な観察記録の山に埋もれて迷っている中にさっさと切り込んで、彼らにかわって明快な論理で本質的原理を見いだしてやるのが自分達の役割だと考える「物理学帝国主義」の信奉者が少なくないが、メンデルはその意味では帝国主義の理想的な実践例であったともいえる。
ブルノはオロミュッツより70キロほど南の、モラビア地方の南端にある町である。チェコの中といっても、プラハまでは200キロもあるが、真南のウィーンまでは110キロしかない。オーストリア・ハンガリー帝国時代には今よりもいっそう、帝国の首都との連絡は密だったに違いない。メンデルは理科の正式教員の免許を取るため、1850年にウィーンに赴いて教員資格試験を受けるが落ちてしまう。だが試験委員長には良い印象を持たれたらしく、ウィーン大学への留学を勧められ、修道院長の理解もあって修道院派遣の学生として2年間の留学を認められた。メンデルは主に物理学を学び、指導教官は(救急車のサイレン音の研究で名高い)ドップラーであった。1853年に留学を終えて修道院に戻ったメンデルは、開設されたばかりの工業学校で実験物理を教えはじめ、その仕事は彼が修道院長になるまでずっと続けられた。帰還3年後の1856年から始められたエンドウ豆を中心とする遺伝の実験が、当時の生物学の仕事としては異様なほど緻密な実験計画と論理的考察に基づいて進められているのは、彼の物理のバックグラウンドが間違いなく影響しているのであろう。1865年に発表された彼の論文が、別刷りを世界中の著名な生物研究者に送りつけるという彼の努力にも関わらず当時ほとんど理解されなかったのは、ある意味では無理もない。物理学者の間には、化学や地学や生物の連中が膨大な観察記録の山に埋もれて迷っている中にさっさと切り込んで、彼らにかわって明快な論理で本質的原理を見いだしてやるのが自分達の役割だと考える「物理学帝国主義」の信奉者が少なくないが、メンデルはその意味では帝国主義の理想的な実践例であったともいえる。
 論文を出して3年後の1868年、メンデルはナップの後を継いで修道院長に選ばれた。カソリックの世界で、修道院長というのは非常に位の高い聖職者である。大学で言えば、学部長よりは偉く、学長に近い。とうぜん雑務も多く、はじめは修道院長の地位を利用してもっと大規模に実験をしようと呑気に考えていたらしいメンデルは、すぐに考えの誤りに気づいたに違いない。自分の論文が世に認められないこと、研究がもはや続けられないことに対する彼の不満をしのばせる言動の記録が、幾つも残っている。だが地域の学校教育の管轄権や修道院への課税を巡っての当局との政治的交渉は、ある意味で彼を燃え立たせるのに十分であったらしく、晩年のメンデルは人間嫌いに陥りつつも、科学からほとんど足を洗ったまま1884年に世を去った。
論文を出して3年後の1868年、メンデルはナップの後を継いで修道院長に選ばれた。カソリックの世界で、修道院長というのは非常に位の高い聖職者である。大学で言えば、学部長よりは偉く、学長に近い。とうぜん雑務も多く、はじめは修道院長の地位を利用してもっと大規模に実験をしようと呑気に考えていたらしいメンデルは、すぐに考えの誤りに気づいたに違いない。自分の論文が世に認められないこと、研究がもはや続けられないことに対する彼の不満をしのばせる言動の記録が、幾つも残っている。だが地域の学校教育の管轄権や修道院への課税を巡っての当局との政治的交渉は、ある意味で彼を燃え立たせるのに十分であったらしく、晩年のメンデルは人間嫌いに陥りつつも、科学からほとんど足を洗ったまま1884年に世を去った。
メンデルの修道院の建物は、現在ではもはや修道院としては機能していないが、ブルノの町のはずれにいまもそのまま残っている(写真6)。メンデルが交配実験をした温室は写真手前の中庭にあったらしいが、整地されていまは跡形もない。門に近い部屋は改装されてメンデル記念館になっている(写真6の1階のいちばん右側のドアが、記念館の入り口)。記念館の前には赤白の花を使って、奥からP(赤白)、F1(赤)、F2(3:1)、F3(9:3:3:1)の遺伝の法則を表現したしゃれた植え込みなどがしつらえてある(写真7)。館内には写真や遺伝学の説明や自筆の実験ノートなどが展示されている(写真8)。だがふつうヨーロッパで記念館といえば、必ずその人の書斎やら机やらが展示してあるものだ。メンデルの部屋は修道院の2階の奥、写真6で木の梢に窓が覆われたあたりにあった。彼の部屋には、しかし中には何も残っていない。
 生物の性質が予め遺伝子に書かれた内容に従って決まってしまうという遺伝学の教えは、曲解すれば個人の努力を否定し、生まれによる差別を正当化することにつながりかねない。社会主義のソ連では、それを避けるため環境条件の変化で生物の遺伝的性質を方向づけ変化させうると説いたルイセンコ(1898-1976)の説が重宝された。また保守的な教会は革命的な立場とはどうも折り合いが悪く、フランス革命の後も、ロシア革命の後も、キリスト教は厳しい弾圧を受けた。第二次大戦後共産圏に組み込まれたチェコスロバキアでも、1949年にはメンデルの修道院は閉鎖されてしまった。また遺伝学のシンボルである彼の足跡は、ルイセンコ主義信奉者の恰好の標的になった。博物館の人によれば、メンデルゆかりの机や椅子などは、そのころの混乱の中で、持ち去られて捨てられてしまったのだそうである。メンデルが再びチェコで正当に評価されるようになったのは、ルイセンコ学派の嵐が去った1960年代後半のことであった。
生物の性質が予め遺伝子に書かれた内容に従って決まってしまうという遺伝学の教えは、曲解すれば個人の努力を否定し、生まれによる差別を正当化することにつながりかねない。社会主義のソ連では、それを避けるため環境条件の変化で生物の遺伝的性質を方向づけ変化させうると説いたルイセンコ(1898-1976)の説が重宝された。また保守的な教会は革命的な立場とはどうも折り合いが悪く、フランス革命の後も、ロシア革命の後も、キリスト教は厳しい弾圧を受けた。第二次大戦後共産圏に組み込まれたチェコスロバキアでも、1949年にはメンデルの修道院は閉鎖されてしまった。また遺伝学のシンボルである彼の足跡は、ルイセンコ主義信奉者の恰好の標的になった。博物館の人によれば、メンデルゆかりの机や椅子などは、そのころの混乱の中で、持ち去られて捨てられてしまったのだそうである。メンデルが再びチェコで正当に評価されるようになったのは、ルイセンコ学派の嵐が去った1960年代後半のことであった。
(因みにルイセンコが広辞苑には載っているのに生物学事典や理化学辞典に載っていないのは、政治や歴史に目をつぶる日本の「学者」の視野の狭さを反映しているように思えて嫌な気分である。)
メンデルがエンドウの実験を始めた動機の一つは、ワインの材料に使うブドウの品種改良に際し、理論的な指針を得るモデル系にする点にあったらしい。彼はエンドウだけでなくブドウを始め多くの植物の育種を手がけていた。第二次大戦のころまでは、これらのブドウはまだ修道院の中庭に植わっていたが、戦後の混乱で引っこ抜かれてしまった。岩波文庫の「雑種植物の研究」(新版)によると、1913年に修道院を訪れた小石川植物園の三好氏が一枝を譲り受け、日本で維持していたブドウが、1989年に再びチェコに送られて、南モラビアの実験圃場に里帰りしたという楽しい話が出ている。1992年の時点では、このブドウはまだ修道院の庭には(少なくとも観光客にも分かる形では)植え替えられていなかった。今ごろ再び訪ねてみれば、日本経由のメンデルのブドウが、記念館の庭に植わっているのであろうか。
(なおこの新版には、メンデルの生地の名前がヒンツィツェというチェコ語でもちゃんと紹介されている。この版をもう少し早く出してくれていれば、オドラウでさまよわずに済んだのに。)
チェコみやげに大量のビールを買い込み、国境の税関で見つからないようトランクの底に隠しこんで、ブルノから南下してウィーンに向かった。巡礼の最後に、ほんとうならウィーン大学をも訪ね、メンデルが学んだ校舎を探してみるべきであったのだが、ウィーンには他に見るものがあまりに多すぎた。美術館巡りに時間が取られ、次の機会にまた探してみようと思ったきり、それから何回はウィーンは訪れたのだが、まだドップラーの研究室の場所探しはやっていない。
シンポジウムお知らせ
山元行動進化プロジェクト・シンポジウム
- 性行動の遺伝解析と進化-
99年8月27日(金)
プログラム
開会前(12:30-13:00) プロジェクトビデオ放映
御挨拶
13:00-13:05 科学技術振興事業団・理事 室田 幹雄
13:05-13:10 プロジェクト・総括責任者 山元 大輔
題目
特別講演 I(13:10-13:40)
Speciation in the Hawaiian Drosophilidae: the role of sexual selection
研究顧問 Kenneth K. Kaneshiro
研究報告
1.キイロショウジョウバエの雌の配偶行動に関するspinster 遺伝子の機能と進化
13:40-14:00 中野 芳朗
2.ハワイのショウジョウバエにみる脳の性的二型とその進化
14:00-14:20 近藤 康弘
3.Genomic organization and characterization of the behavior gene fruitless (fru) in the Hawaiian species Drosophila heteroneura and D. silvestris and conservation of the fru BTB protein-protein-binding domain throughout evolution
14:20-14:40 Terence Davis
休憩(14:40-15:10)
4.Dual tagging gene-trap in Drosophila:a new efficient technique for func tional genomics
15:10-15:40 Tamas Lukacsovich 粟野 若枝
5.交尾における生殖器の連結および解除に異常を示すキイロショウジョウバエの変異体、lingerer の解析
15:40-16:10 国吉 久人
6.fickle 変異体の解析から同定されたショウジョウバエ Bruton's tyrosine kinase (Btk) ホモログの構造と機能
16:10-16:40 従二 直人
7.雄の性指向性と雄特異的筋肉の形成を支配する satori (fruitless) 遺伝子の解析
16:40-17:10 高橋 邦明 青木 一恵
特別講演 II (17:10-17:40)
Chemical communication in Drosophila
客員研究員 Jean-Marc Jallon
研究の総括(17:40-18:00)
性行動の遺伝解析と進化
プロジェクト・総括責任者 山元 大輔
懇親会(18:00-20:00)
場所:東京ガーデンパレス(東京都文京区湯島1-7-5)
・JR御茶ノ水駅下車徒歩5分
・地下鉄丸ノ内線御茶ノ水駅下車徒歩5分
・地下鉄千代田線新御茶ノ水駅下車徒歩5分
いずれも聖橋方面より
参加費:無料
参加申込:事前申込(先着250名、7月25日まで)
懇親会:2000円、事前申込(7月25日まで)
申込先:〒194-8511 東京都町田市南大谷11号 三菱化学生命科学研究所内
科学技術振興事業団 ERATO 山元行動進化プロジェクト
TEL:042-721-2334 FAX:042-721-2850
E-mail:snishi@fly.erato.jst.go.jp
「日本ショウジョウバエ研究会」会則
1. 本会を「日本ショウジョウバエ研究会」(JDRC: Japanese Drosophila Research C
onference)と称する。
2. 本会はショウジョウバエを利用した研究を行う人およびショウジョウバエ研究に
興味を持つ人を広く結集し、相互の情報交換と研究討議を行う場を提供する。
3. 上記の趣旨に賛同する者は誰でも会員になれる。
4. 本会には代表1人を含め世話役4人を置く。
5. 代表は本会運営の責任者であり、また研究集会の計画・実行に責任を持つ。代表
は研究集会が開かれるごとに交代する。
6. 代表以外の世話役のうち、1人は「ショウジョウバエ通信」の編集を担当し、全
員に必要な情報を伝達する。
7. 他の1人が事務局を分担し、名簿の管理、通信の発送、金銭の出納を行う。
8. 世話役は会員全体のために、緊密に連絡し広い視野で活動する。
9. 世話役は2人ずつ2年毎に交代し連続4年勤める。4年を越えるのは認めない。世
話役を辞めたのち4年間は再び世話役になることができない。
10. 新しい世話役は地域、研究分野等のバランスを考慮して現世話役が決定する。会
員は世話役に候補者を推薦することができる。
11. 世話役の交代は研究集会時に行い、出席者の承認を得る。
12. 新しい世話役に不満がある者は代わりの候補を立てて選挙を要求できる。
13. 会の運営費が不足してきた時には、これを徴収することがある。この場合、研究
室を主管する立場にある者が多くを負担するものとする。
14.この規定に無かったり、詳しく決めてない事態が発生した場合の処理は世話役に
一任する。
補則
1. 入会するには事務局へ申込む。研究会名簿に登録され「ショウジョウバエ通信」
を受け取ることができる。同時にJfly mailing listにも参加されるようお勧めします。
2. Jfly は伊藤啓氏のボランティアによって運営されています。ショウジョウバエ研
究会の活動ではありませんが、良好な協力関係のもとに、情報伝達等に利用させても
らっています。
3. 何らかの事情で任期中に世話役の交代が必要になる場合の手続きは14 項を適用し
て対処する。
(以上)
_______________________________________________________________
日本ショウジョウバエ研究会(世話役)
西田 育巧(代表):〒464-8602 名古屋市千種区不老町 名古屋大学
大学院理学研究科 生命理学専攻 発生生物学研究グループ Tel: 052-789-2472
Fax: 052-789-2511 E-mail: nishida@bio.nagoya-u.ac.jp
林 茂生:〒411-8540 三島市谷田1111 国立遺伝学研究所 系統生物研究センター
無脊椎動物遺伝研究室 Tel: 0559-81-6823 Fax: 0559-81-6825
E-mail: shayashi@lab.nig.ac.jp
丸尾 文昭(事務局): 〒305-8572 つくば市天王台1-1-1 筑波大学生物科学系
Tel: 0298-53-4909 Fax: 0298-53-6669 E-mail: maru@biol.tsukuba.ac.jp
木村 賢一(通信):〒068-8642 岩見沢市緑が丘2丁目 北海道教育大学岩見沢校
生物研究室 Tel: 0126-32-0341 Fax: 0126-32-0255
E-mail: kimura@iwa.hokkyodai.ac.jp
ホームページ:http://www.biol.tsukuba.ac.jp/~maru/JDRC/
Jfly:伊藤 啓 〒444-8585 愛知県 岡崎市明大寺町 字西郷中 38番地
岡崎国立共同研究機構 基礎生物学研究所 細胞増殖部門 Tel: 0564-55-7532
Fax: 0564-55-7533 E-mail: iitokei@nibb.ac.jp
Jfly サーバー: http://jfly.nibb.ac.jp/index_j.html
_______________________________________________________________
木村賢一
北海道教育大学岩見沢校 生物研究室
〒068-8642 岩見沢市緑が丘2丁目
tel: 0126-32-0341
fax: 0126-32-0255
E-mail: kimura@iwa.hokkyodai.ac.jp
 2時間ほど走り回り、本当の村の場所は分からなかったけれど、まあこのような景色を見ながらメンデルは育ったのだろうということだけで満足して帰ろうかと思いだしたころ、小さな食堂を見つけた。昼下がりで、近所の人らしいのが15人ほど新聞を読んだり世間話をしたりしている。片端から聞いてみれば誰か一人くらいは知っているかもしれないと考え、店の中に入ってみた。たいへん運の良いことに、客の中にドイツ語が分かり、しかもタクシー運転手をしているという願ってもない人がいた。ああ、それなら多分ヒンツィーチェ Hyncice だ。オドラウのちょっと南だよ。と差し出した地図に印をつけてくれた。
2時間ほど走り回り、本当の村の場所は分からなかったけれど、まあこのような景色を見ながらメンデルは育ったのだろうということだけで満足して帰ろうかと思いだしたころ、小さな食堂を見つけた。昼下がりで、近所の人らしいのが15人ほど新聞を読んだり世間話をしたりしている。片端から聞いてみれば誰か一人くらいは知っているかもしれないと考え、店の中に入ってみた。たいへん運の良いことに、客の中にドイツ語が分かり、しかもタクシー運転手をしているという願ってもない人がいた。ああ、それなら多分ヒンツィーチェ Hyncice だ。オドラウのちょっと南だよ。と差し出した地図に印をつけてくれた。 南下してオドラウに戻り、もう2キロほど街道を南下してから、それて脇道に入る。さらに2キロほど行くと、「オドラウ郡ヒンツィーチェ」の看板が道端に見つかった。嬉しさのあまり思わず車を停めて、しばし感慨に浸る。ヒンツィーチェの村は、ゆるやかな起伏の畑地のなかに、ちょっと盛り上がった小さな丘の上にあった(写真1)。一本道がいったん坂を下り、村に続く丘のふもとに達したあたりの、上り坂になる少し手前の村の入口に、くすんだ茶色が普通のこのあたりの民家と異なり、やけに立派な白塗りの小屋があった(写真2)。屋根には豪勢な破風がつけられ、ドイツ語の花文字が書かれた石版が飾られている(写真3)。読んで見ると:
南下してオドラウに戻り、もう2キロほど街道を南下してから、それて脇道に入る。さらに2キロほど行くと、「オドラウ郡ヒンツィーチェ」の看板が道端に見つかった。嬉しさのあまり思わず車を停めて、しばし感慨に浸る。ヒンツィーチェの村は、ゆるやかな起伏の畑地のなかに、ちょっと盛り上がった小さな丘の上にあった(写真1)。一本道がいったん坂を下り、村に続く丘のふもとに達したあたりの、上り坂になる少し手前の村の入口に、くすんだ茶色が普通のこのあたりの民家と異なり、やけに立派な白塗りの小屋があった(写真2)。屋根には豪勢な破風がつけられ、ドイツ語の花文字が書かれた石版が飾られている(写真3)。読んで見ると: 日本でも村の入口には、よく「○○消防団」と書かれた小さな倉庫のような建物が建っている。村との位置関係といい建物の大きさといい、ちょうど同じである。例の岩波文庫の「メンデル小傳」には、「彼がいよいよ有名になった時に、新しい消防署の建立が、彼を記念する機會として用ひられて、記念碑をその建物に掲げて表彰した」とある。消防団にしては屋根だけがやけに立派なのが、面白い。開いていた扉から中を覗くと、荒れ果てた物置きになっていてほとんど使われていない様子であった。
日本でも村の入口には、よく「○○消防団」と書かれた小さな倉庫のような建物が建っている。村との位置関係といい建物の大きさといい、ちょうど同じである。例の岩波文庫の「メンデル小傳」には、「彼がいよいよ有名になった時に、新しい消防署の建立が、彼を記念する機會として用ひられて、記念碑をその建物に掲げて表彰した」とある。消防団にしては屋根だけがやけに立派なのが、面白い。開いていた扉から中を覗くと、荒れ果てた物置きになっていてほとんど使われていない様子であった。 丘の上の村へ続くカーブした坂をのぼり切ると、まっすぐな道の両側に数十軒ほどの家が並ぶヒンツィーチェの村に出る。家並みの真ん中へんには道の片側に教会が立っていて、そのへんが村の中心らしい。メンデルの家はこの村の58番地、58番地と言えば坂をのぼり切ってすぐ、家並みの一番はずれのあたりである。その位置には茶色い三角屋根の家が建っている。道路に面した壁の中程には、小さなプレートが埋め込んである。こんどはチェコ語なので何と書いてあるのかよく分からないが、Mendel という文字は判読できる(写真4)。これが生家だと信じてよさそうだ。180年前の家がそのまま残っているのかどうかは確かめられなかったが、チェコはほとんど戦災にあっていないから、その確率は高い。
丘の上の村へ続くカーブした坂をのぼり切ると、まっすぐな道の両側に数十軒ほどの家が並ぶヒンツィーチェの村に出る。家並みの真ん中へんには道の片側に教会が立っていて、そのへんが村の中心らしい。メンデルの家はこの村の58番地、58番地と言えば坂をのぼり切ってすぐ、家並みの一番はずれのあたりである。その位置には茶色い三角屋根の家が建っている。道路に面した壁の中程には、小さなプレートが埋め込んである。こんどはチェコ語なので何と書いてあるのかよく分からないが、Mendel という文字は判読できる(写真4)。これが生家だと信じてよさそうだ。180年前の家がそのまま残っているのかどうかは確かめられなかったが、チェコはほとんど戦災にあっていないから、その確率は高い。 成績が良かったメンデルは、学校の先生の薦めもあり、ギムナジウムを経て大きな町の高校(日本の旧制高校に当たる)へ進学することができた。北部モラビア随一の町は、当時はオロミュッツ、現在はオロモウツと呼ばれる、このあたりの司教座聖堂もある町である。(ちなみにベートーベンの「戴冠ミサ曲」は、彼の友人がウィーンからオロミュッツの司教になって赴任した際に書いた曲である。)メンデルに従い、オロモウツへ向かう。ヒンチーチェからは約50キロ、当時はまる1日以上の旅程だっただろう。当然メンデルは寄宿生活を送っていたはずだ。父の病気で仕送りも滞りがちになり、しかも自身も3回も病気を患って、必ずしも明るく楽しい高校生活ではなかったようである。
成績が良かったメンデルは、学校の先生の薦めもあり、ギムナジウムを経て大きな町の高校(日本の旧制高校に当たる)へ進学することができた。北部モラビア随一の町は、当時はオロミュッツ、現在はオロモウツと呼ばれる、このあたりの司教座聖堂もある町である。(ちなみにベートーベンの「戴冠ミサ曲」は、彼の友人がウィーンからオロミュッツの司教になって赴任した際に書いた曲である。)メンデルに従い、オロモウツへ向かう。ヒンチーチェからは約50キロ、当時はまる1日以上の旅程だっただろう。当然メンデルは寄宿生活を送っていたはずだ。父の病気で仕送りも滞りがちになり、しかも自身も3回も病気を患って、必ずしも明るく楽しい高校生活ではなかったようである。 ブルノはオロミュッツより70キロほど南の、モラビア地方の南端にある町である。チェコの中といっても、プラハまでは200キロもあるが、真南のウィーンまでは110キロしかない。オーストリア・ハンガリー帝国時代には今よりもいっそう、帝国の首都との連絡は密だったに違いない。メンデルは理科の正式教員の免許を取るため、1850年にウィーンに赴いて教員資格試験を受けるが落ちてしまう。だが試験委員長には良い印象を持たれたらしく、ウィーン大学への留学を勧められ、修道院長の理解もあって修道院派遣の学生として2年間の留学を認められた。メンデルは主に物理学を学び、指導教官は(救急車のサイレン音の研究で名高い)ドップラーであった。1853年に留学を終えて修道院に戻ったメンデルは、開設されたばかりの工業学校で実験物理を教えはじめ、その仕事は彼が修道院長になるまでずっと続けられた。帰還3年後の1856年から始められたエンドウ豆を中心とする遺伝の実験が、当時の生物学の仕事としては異様なほど緻密な実験計画と論理的考察に基づいて進められているのは、彼の物理のバックグラウンドが間違いなく影響しているのであろう。1865年に発表された彼の論文が、別刷りを世界中の著名な生物研究者に送りつけるという彼の努力にも関わらず当時ほとんど理解されなかったのは、ある意味では無理もない。物理学者の間には、化学や地学や生物の連中が膨大な観察記録の山に埋もれて迷っている中にさっさと切り込んで、彼らにかわって明快な論理で本質的原理を見いだしてやるのが自分達の役割だと考える「物理学帝国主義」の信奉者が少なくないが、メンデルはその意味では帝国主義の理想的な実践例であったともいえる。
ブルノはオロミュッツより70キロほど南の、モラビア地方の南端にある町である。チェコの中といっても、プラハまでは200キロもあるが、真南のウィーンまでは110キロしかない。オーストリア・ハンガリー帝国時代には今よりもいっそう、帝国の首都との連絡は密だったに違いない。メンデルは理科の正式教員の免許を取るため、1850年にウィーンに赴いて教員資格試験を受けるが落ちてしまう。だが試験委員長には良い印象を持たれたらしく、ウィーン大学への留学を勧められ、修道院長の理解もあって修道院派遣の学生として2年間の留学を認められた。メンデルは主に物理学を学び、指導教官は(救急車のサイレン音の研究で名高い)ドップラーであった。1853年に留学を終えて修道院に戻ったメンデルは、開設されたばかりの工業学校で実験物理を教えはじめ、その仕事は彼が修道院長になるまでずっと続けられた。帰還3年後の1856年から始められたエンドウ豆を中心とする遺伝の実験が、当時の生物学の仕事としては異様なほど緻密な実験計画と論理的考察に基づいて進められているのは、彼の物理のバックグラウンドが間違いなく影響しているのであろう。1865年に発表された彼の論文が、別刷りを世界中の著名な生物研究者に送りつけるという彼の努力にも関わらず当時ほとんど理解されなかったのは、ある意味では無理もない。物理学者の間には、化学や地学や生物の連中が膨大な観察記録の山に埋もれて迷っている中にさっさと切り込んで、彼らにかわって明快な論理で本質的原理を見いだしてやるのが自分達の役割だと考える「物理学帝国主義」の信奉者が少なくないが、メンデルはその意味では帝国主義の理想的な実践例であったともいえる。 論文を出して3年後の1868年、メンデルはナップの後を継いで修道院長に選ばれた。カソリックの世界で、修道院長というのは非常に位の高い聖職者である。大学で言えば、学部長よりは偉く、学長に近い。とうぜん雑務も多く、はじめは修道院長の地位を利用してもっと大規模に実験をしようと呑気に考えていたらしいメンデルは、すぐに考えの誤りに気づいたに違いない。自分の論文が世に認められないこと、研究がもはや続けられないことに対する彼の不満をしのばせる言動の記録が、幾つも残っている。だが地域の学校教育の管轄権や修道院への課税を巡っての当局との政治的交渉は、ある意味で彼を燃え立たせるのに十分であったらしく、晩年のメンデルは人間嫌いに陥りつつも、科学からほとんど足を洗ったまま1884年に世を去った。
論文を出して3年後の1868年、メンデルはナップの後を継いで修道院長に選ばれた。カソリックの世界で、修道院長というのは非常に位の高い聖職者である。大学で言えば、学部長よりは偉く、学長に近い。とうぜん雑務も多く、はじめは修道院長の地位を利用してもっと大規模に実験をしようと呑気に考えていたらしいメンデルは、すぐに考えの誤りに気づいたに違いない。自分の論文が世に認められないこと、研究がもはや続けられないことに対する彼の不満をしのばせる言動の記録が、幾つも残っている。だが地域の学校教育の管轄権や修道院への課税を巡っての当局との政治的交渉は、ある意味で彼を燃え立たせるのに十分であったらしく、晩年のメンデルは人間嫌いに陥りつつも、科学からほとんど足を洗ったまま1884年に世を去った。 生物の性質が予め遺伝子に書かれた内容に従って決まってしまうという遺伝学の教えは、曲解すれば個人の努力を否定し、生まれによる差別を正当化することにつながりかねない。社会主義のソ連では、それを避けるため環境条件の変化で生物の遺伝的性質を方向づけ変化させうると説いたルイセンコ(1898-1976)の説が重宝された。また保守的な教会は革命的な立場とはどうも折り合いが悪く、フランス革命の後も、ロシア革命の後も、キリスト教は厳しい弾圧を受けた。第二次大戦後共産圏に組み込まれたチェコスロバキアでも、1949年にはメンデルの修道院は閉鎖されてしまった。また遺伝学のシンボルである彼の足跡は、ルイセンコ主義信奉者の恰好の標的になった。博物館の人によれば、メンデルゆかりの机や椅子などは、そのころの混乱の中で、持ち去られて捨てられてしまったのだそうである。メンデルが再びチェコで正当に評価されるようになったのは、ルイセンコ学派の嵐が去った1960年代後半のことであった。
生物の性質が予め遺伝子に書かれた内容に従って決まってしまうという遺伝学の教えは、曲解すれば個人の努力を否定し、生まれによる差別を正当化することにつながりかねない。社会主義のソ連では、それを避けるため環境条件の変化で生物の遺伝的性質を方向づけ変化させうると説いたルイセンコ(1898-1976)の説が重宝された。また保守的な教会は革命的な立場とはどうも折り合いが悪く、フランス革命の後も、ロシア革命の後も、キリスト教は厳しい弾圧を受けた。第二次大戦後共産圏に組み込まれたチェコスロバキアでも、1949年にはメンデルの修道院は閉鎖されてしまった。また遺伝学のシンボルである彼の足跡は、ルイセンコ主義信奉者の恰好の標的になった。博物館の人によれば、メンデルゆかりの机や椅子などは、そのころの混乱の中で、持ち去られて捨てられてしまったのだそうである。メンデルが再びチェコで正当に評価されるようになったのは、ルイセンコ学派の嵐が去った1960年代後半のことであった。