|
|
|
|
|

|
当研究室では、キイロショウジョウバエ(Drosophila melanogaster)を利用して、脳の構造・機能・発生過程の解析を行なっています。 脳構造の分子解剖学的な解析
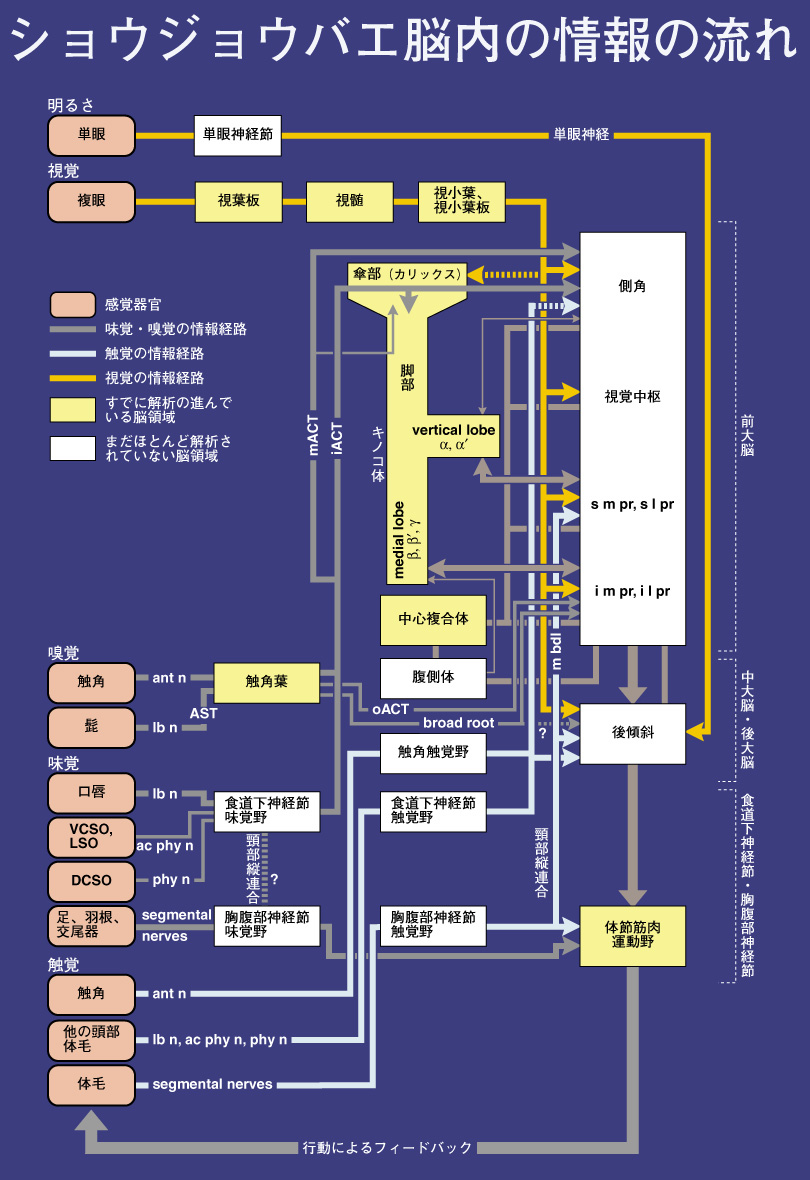
脳内の機能部位の解析
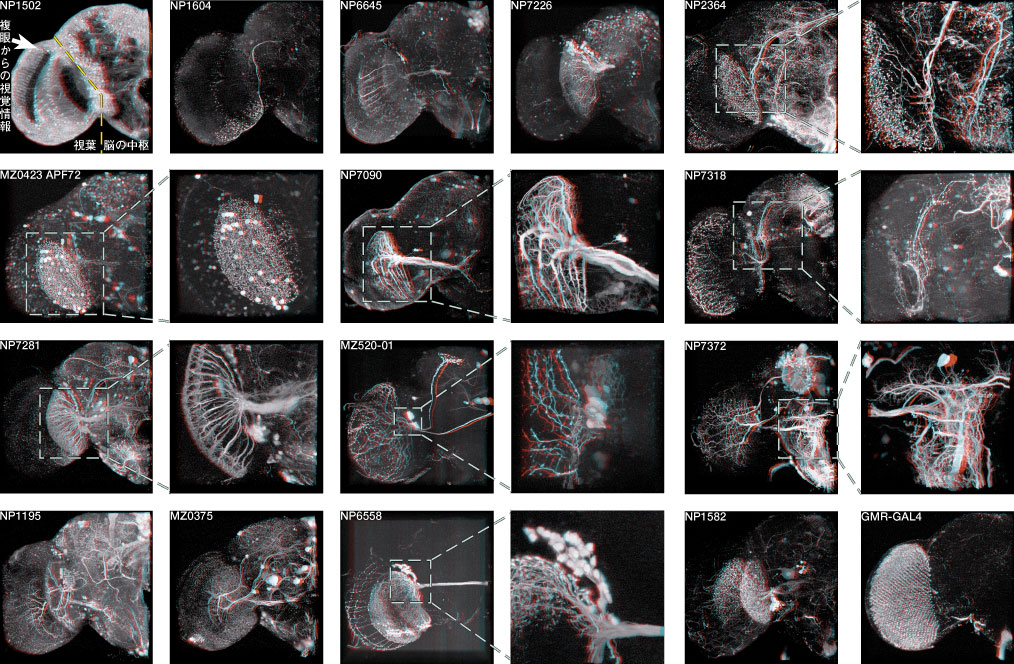
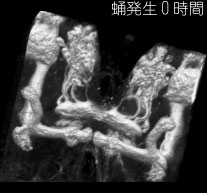
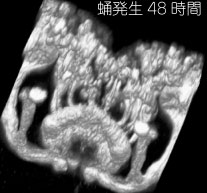
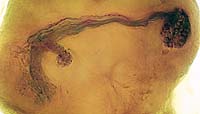
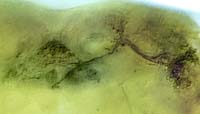
神経回路の発生過程の解析設計図をシリコンに焼き付ければ完成するコンピューターと違い、脳では個々の細胞が、発生過程で自ら線維を伸ばしながら、神経回路網を作っていきます。この回路形成のメカニズムを探るには、まず神経線維がいつ、どのようにして伸びてくるかをきちんと観察することが、大前提になります。個々の細胞の線維を見分けるのが難しいため、従来このような観察は、網膜視蓋投射のような太い軸索束や、中枢から筋肉へ伸びる末梢神経に着目した研究に限られていました。しかし特定の神経細胞をラベルした GAL4 エンハンサートラップ系統を用いると、多数の線維が錯綜した連合野における細かな介在神経についても、成虫から遡ってさまざまな発生段階での標本を作り、回路形成の過程を調べることが出来ます。本研究室では、成虫で神経細胞を同定すると同時に、その細胞の線維伸長過程を発生段階を追って解析し、そこに関与する分子メカニズムを研究しています。
脳回路網と細胞系譜の関係の解析
脳のバイオインフォマティクスの研究脳の情報学的研究では、ボトムアップ型とトップダウン型という2つのアプローチがよく提唱されます。神経解剖学や神経生理学によって単一細胞レベルから神経回路の構造と機能を調べてゆくボトムアップ型研究に対し、神経回路の実体をいったん離れ、回路が持つべき構造や果たすべき機能をコンピューターシミュレーションなどによって情報科学的に推理するのが、トップダウン型研究です。脳研究の進展にともなって両手法は徐々に融合することが期待されてきました。しかし両手法が本格的に始まってすでに数十年経つにも関わらず、融合の兆しはなかなか見えません。極度に単純化された仮想回路でなく、現実の神経回路に基づいた脳機能を再現するようなコンピューターシミュレーションは、未だ実現にはほど遠い状況です。原因の一つは、21世紀初頭という現時点において、生物学の解析技術でボトムアップ型研究で調べることが可能な神経回路の情報はきわめて断片的で限られているのに対し、トップダウン型研究では認知・学習など多数の脳領域の複雑な協調処理が不可欠な、極めて高度な脳機能の解析に大きな関心が持たれていて、両者の間の乖離が大きすぎる点にあります。 神経回路が比較的小さく単純で、高度な脳機能のレベルにも限りがあるショウジョウバエの脳は、ボトムアップ型研究とトップダウン型研究を現実的に融合しうる、数少ない実験系です。本研究室では、比較的単純なショウジョウバエ脳の機能をコンピューター上に再現することを長期目標としています。しかし現時点では、解析された脳神経回路の情報をどのようにコンピューター内に格納するかさえ、定まった方式はありません。まず第一歩として、顕微鏡断層撮影像から神経の細胞位置や回路のトポロジー構造を抽出し、データベース化する手法の開発を進めています。 バックグラウンドなぜ地味で記載的な解剖学なのか。なぜショウジョウバエなのか。補足説明します。最近の論文Kamikouchi, A., Inagaki, H. K., Effertz, T., Fiala, A., Hendrich, O., Gopfert, M. C. and Ito, K. (2009) The neural basis of Drosophila gravity sensing and hearing. Nature (Article), 458, 165-171.Yorozu, S., Wong, A., Fischer, B. J., Dankert, H., Kernan, M. J., Kamikouchi, A., Ito, K., Anderson, D.J. (2009) Distinct sensory representations of wind and courtship song in the Drosophila brain. Nature (Letter), 458, 201-205. Okada, R., Awasaki, T. and Ito, K. (2009) GABA-mediated neural connections in the Drosophila antennal lobe. J Comp Neurol, 514, 74-91. Kato, K., Awasaki, T. and Ito, K. (2009) Neuronal programmed cell death induces glial cell division in the adult Drosophila brain. Development, 136, 51-9. Tanaka, N. K., Tanimoto, H., and Ito, K. (2008) Neuronal assemblies of the Drosohila mushroom body. J Comp Neurol. (508) 711-755 Ito, K. and Awasaki, T. (2008) Clonal unit architecture of the adult fly brain. Textbook “Brain development in Drosophila” pp.137-158, Landes Bioscience/Springer, USA Awasaki, T., Lai, S. L., Ito, K. and Lee, T. (2008) Organization and post-embryonic development of glial cells in the adult central brain of Drosophila. J Neurosci, 28, 13742-13753. Silbering, A. F., Okada, R., Ito, K. and Galizia, C. G. (2008) Odor information processing in the Drosophila antennal lobe: Anything goes? J Neurosci 28, 13075-87. Busch, S., Selcho, M., Ito, K. and Tanimoto, H. (2008) A map of octopaminergic neurons in the Drosophila brain. J Comp Neurol, 513, 643-667. Endo, K., Aoki, T., Yoda, Y., Kimura, K., Hama, C. (2007) Notch signal organizes the Drosophila olfactory circuitry by diversifying the sensory neuronal lineages. Nat. Neurosci. (10) 153-60 Sachse, S., Rueckert, E., Keller, A., Okada, R., Tanaka, N. K., Ito, K. and Vosshal, L. B. (2007) Activity-dependent plasticity in an olfactory circuit. Neuron (56) 838-50 Kamikouchi A, Shimada, T. and ITO, K. (2006) Comprehensive classification of the auditory sensory projections in the brain of the fruit fly Drosophila melanogaster. J. Comp. Neurol. (499) 317-356, 2006. Otsuna, H. & Ito, K. (2006) Systematic Analysis of the Visual projection neurons of Drosophila melanogaster - I: Lobula-specific pathways. J. Comp. Neurol. (497) 928-958. Awasaki, T., Tatsumi, R., Takahashi, K., Arai, K., Nakanishi, Y., Ueda, R., and Ito, K. (2006) Essential role of the apoptotic cell engulfment genes draper and ced-6 in programmed axon pruning during Drosophila metamorphosis. Neuron (50) 855-867 Liu. G., Seiler, H., Wen, A., Zars, T., Ito,. K., Wolf, R., Heisenberg, M. & Liu, L. (2006). Distinct memory traces for two visual features in the Drosophila brain. Nature (439) 551-556. 以前の主な論文 Tanaka, N. K., Awasaki, T., Shimada, T., and Ito, K. (2004). Integration of chemosensory pathways in the Drosophila second-order olfactory centers. Curr. Biol. (14) 449-457 Awasaki, T., and Ito, K. (2004). Engulfing action of glial cells is required for programmed axon pruning during Drosophila metamorphosis. Curr. Biol. (14) 668-677 Kido, A., and Ito, K. (2002). Mushroom bodies are not required for courtship behavior by normal and sexually mosaic Drosophila. J Neurobiol (52) 302-311. Verkhusha, V. V., Otsuna, H., Awasaki, T., Oda, H., Tsukita, S. and Ito, K. (2001). An enhanced mutant of red-fluorescent protein DsRed for double labeling and developmental timer of neural fiber bundle formation. J. Biol. Chem. (276) 29621-29624 Awasaki, T., Saito, M., Sone, M., Suzuki, E., Sakai, R., Ito, K., Hama, C. (2000) The Drosophilla Trio plays an essential role in patterning of axons by regulating their directional extention . Neuron (26) 119-131 Ito, K., Suzuki, K., Estes, P., Ramaswami, M., Yamamoto, D. and Strausfeld, N. J. (1998). The organization of extrinsic neurons and their implications regarding the functional roles of the mushroom bodies in Drosophila melanogaster Meigen. Learning and Memory (5) 52-77. Ito, K., Awano, W., Suzuki, K., Hiromi, Y. and Yamamoto, D. (1997). The Drosophila mushroom body is a quadruple structure of clonal units each of which contains a virtually identical set of neurones and glial cells. Development (124) 761-771. Ito, K., Urban, J. and Technau, G. M. (1995). Distribution, classification, and development of Drosophila glial cells in the late embryonic and early larval ventral nerve cord. Roux's Arch. Dev. Biol. (204) 284-307. Ito, K. and Hotta, Y. (1992). Proliferation pattern of postembryonic neuroblasts in the brain of Drosophila melanogaster. Dev. Biol. (149) 134-148. 総説: 伊藤啓 (2005). ショウジョウバエにおける蛍光イメージング実験. バイオテクノロジージャーナル (5) 477-485. Ito, K., Okada, R., Tanaka, N. K. and Awasaki, T. (2003). Cautionary observations on preparing and interpreting brain images using molecular biology-based staining techniques. Microsc Res Tech(62) 170-186. 伊藤啓 (2000). ショウジョウバエを使って脳の回路図をつくる. 生物物理 40, 179-184. Strausfeld, N. J., Hansen, L., Li, Y., Gomez, R. S. and Ito, K. (1998). Evolution, discovery, and interpretations of Arthropod mushroom bodies. Learning and Memory (5) 11-37. リンク・JT 生命誌館ホームページ「Scientist Library」の関連ページへ(新版)・JT 生命誌館ホームページ「Scientist Library」の関連ページへ(旧版) ・Scientist Library ホームページへ |