



|
|
|
|
|

|
昆虫脳の脳構造を定義する枠組みを完成Ito.K., Shinomiya, K., Ito, M., Armstrong, D., Boyan, G., Hartenstein, V., Harzsch, S., Heisenberg, M., Homberg, U., Jenett, A., Keshishian. H., Restifo, L., Rossler, W., Julie Simpson, J., Strausfeld, N. J., Strauss, R., and Vosshall, L.B; The Insect Brain Name Working Group.A systematic nomenclature for the insect brain. Neuron, 81, 755-765, 2014.
全脳レベルの神経回路解析が他の生物にさきがけて進んでいる、ショウジョウバエをはじめとする昆虫脳でも、この問題は重要である。そのため、5年以上の歳月をかけて、4ケ国の15研究室からなる国際チームをまとめ、今後の脳研究の共通基盤となる「昆虫脳のすべての脳構造を定義する体系の枠組み」をショウジョウバエの脳を基準として作成し、公表した。この枠組みでは最新の脳研究成果を総合して脳全体を43の領域に区分し、境界と名前を定義した。また、混乱やばらつきがある用語37種類について解決策を定めた。新たな枠組みは、すでに各国の大規模脳データベースで脳の場所を記述する基準として採用されており、今後の昆虫脳研究のスタンダードとなる。 細胞系譜に基づくショウジョウバエ脳全神経投射回路のマップIto, M., Masuda, N., Shinomiya, K., Endo, K., and Ito, K.Systematic analysis of neural projections reveals clonal composition of the Drosophila brain. Curr. Biol, 23, 644-655, 2013.
ショウジョウバエ嗅覚一次中枢の網羅的神経投射マップTanaka, N. K., Endo, K. and Ito, K.The organization of antennal lobe-associated neurons in the adult Drosophila melanogaster brain. J Comp Neurol, 520, 4067-4130, 2012.
ショウジョウバエ脳神経の包括的画像データベースShinomiya, K., Matsuda, K., Oishi, T., Otsuna, H., and Ito, K.Flybrain Neuron Database, a com-prehensive database system of the Drosophila brain neurons. J Comp Neurol, 519, 807-833, 2011. 脳にある様々な神経の形状や投射パターンに関する記述は、多数の文献に、異なった形で掲載されており、情報を一望して比較することが難しい。そこで既知のすべての神経を網羅して、分かっている情報を統一的な形で整理した初のデータベースを作成した。神経に関する情報、脳の領域に関する情報、特定の神経をラベルする分子マーカーや発現誘導系統に関する情報の、3つのデータベースをリンクさせて相互に検索可能にすることによって、膨大な情報を整理して概観できるようにした。さらに、これらの情報を利用して、いくつもの論文に散在した情報をまとめることによって個々の論文では解析できていなかった神経回路ネットワークの特徴を明らかにし、情報を集約して比較することの有用性を示した。 ショウジョウバエ味覚中枢の初の体系的神経投射マップMiyazaki, T. and Ito, K.Neural architecture of the primary gustatory center of Drosophila melano-gaster visualized with GAL4 and LexA enhancer-trap systems. J Comp Neurol, 518, 4147-4181, 2010.
ショウジョウバエ嗅覚一次中枢のGABA抑制性神経回路Okada, R., Awasaki, T. and Ito, K.GABA-mediated neural connections in the Drosophila antennal lobe. J Comp Neurol, 514, 74-91, 2009
本研究と並行して多数の共同研究を行い (Tanaka, Ito, Stopfer. J Neurosci (29) 8595-603, 2009; Das, Sen, Lichtneckert, Okada, Ito, Rodrigues, Reichert, Neural Development (3), 33, 2008; Sachse, Rueckert, Keller, Okada, Tanaka, Ito, Vosshal, Neuron (56) 838-50, 2007)、この結果新たに発見した2種類の抑制性局所神経の発生過程や、匂い情報のコードと反応可塑性に果たす特徴的な機能の違いについて、一挙に理解が促進された。 ショウジョウバエの重力感覚と聴覚の神経基盤Kamikouchi, A., Inagaki, H. K., Effertz, T., Fiala, A., Hendrich, O., Gopfert, M. C. and Ito, K.The neural basis of Drosophila gravity sensing and hearing. Nature (Article), 458, 165-171, 2009.
まず、それぞれのジョンストン器官神経グループで、細胞内カルシウム濃度に依存して蛍光強度が変化するタンパクを発現させ、触角を電気的に動かしながら神経の活動をイメージング解析した。その結果、一次中枢の5つの領域のうちのゾーンCに投射する神経とゾーンEに投射する神経は触角の定常的な変位に、ゾーンAに投射する神経とゾーンBに投射する神経は触角の振動に、それぞれ特異的に反応することが分かった。次に、シナプス小胞の放出を阻害する毒素を発現させて特定の神経の機能を特異的に阻害する実験を行うと、ゾーンCとEに投射する変位検知神経を阻害すると重力を検知する行動実験の反応が特異的に消失し、求愛の羽音を検知する行動実験には影響がないのに対し、ゾーンBに投射する振動検知神経を阻害すると重力への反応には影響がなく、音への反応が特異的に消失することが分かった。また脳内で感覚神経からの情報を受け取る二次神経の構造を解析すると、ハエの重力中枢と音中枢の神経回路構造は、ヒトの脳で重力と音の情報を処理する前庭神経核・蝸牛神経核と、それぞれ高い類似性を持っていた(図)。 これまで視覚・嗅覚・味覚について、哺乳類と昆虫の脳で情報処理回路が類似していることが知られていた。今回の研究で聴覚や重力感覚についても高い類似性が明らかになり、6億年以上前に分かれて別個に進化してきた2つの脳システムの構築原理の理解に新たな光を投じた。 神経細胞死はショウジョウバエ成虫脳のグリア増殖を惹き起こすKato, K., Awasaki, T. and Ito, K.Neuronal programmed cell death induces glial cell division in the adult Drosophila brain. Development, 136, 51-59, 2009.
ショウジョウバエ成虫の脳では、羽化直後の数日間だけ、脳の特定の数カ所で神経のプログラム細胞死が観察される。脳内の細胞増殖をハエの寿命の全期間にわたって体系的に調べた結果、神経幹細胞の成体期の増殖はいっさい見られないかわり、プログラム細胞死と同じ時期に同じ場所で、細胞の増殖が観察された(図の上段)。細胞の形態や分子マーカーの発現パターンから、これらの増殖細胞はグリア細胞だった。グリア増殖は神経のプログラム細胞死よりわずかに遅れて始まり、神経細胞死が見られなくなってしばらくすると終了する。また、プログラム細胞死の抑制遺伝子を神経細胞で発現させて細胞死が起きなくすると、グリア増殖は観察されなくなる。すなわちグリアの増殖は、神経のプログラム細胞死に呼応して起きている。 また、従来外傷などによる神経損傷に対してグリアが増殖する例が哺乳類で知られていたが、ハエの脳に投射する感覚神経を切断したり、脳に細い針を刺したりして神経損傷を起こさせると(図の下段)、損傷部位のまわりに特異的なグリア増殖が観察された。 プログラム細胞死に関与するTNFスーパーファミリーリガンドであるeiger遺伝子の変異体では、神経損傷とプログラム神経細胞死の両方に対して、グリアの増殖が著しく抑制される。この状態でグリア細胞だけに特異的にeiger遺伝子を発現させると、グリア増殖が回復する。従ってグリアでのeigerの機能が、神経損傷とプログラム神経細胞死への対応の両方に必要な、共通の機構であることが分かった。またグリア増殖が抑制されると、神経の細胞死は通常より増加した。つまりグリア増殖は、神経細胞死が周囲に与える影響を抑える効果を持つ。 面白いことに、グリアが増殖するのは羽化後1週間程度までの若い成虫だけで、年取った個体では脳に損傷を与えてもグリアの増殖は見られない。グリア増殖が起きる期間はハエの脳の嗅覚中枢に高い可塑性が見られる期間と一致しており、ハエ成虫の脳機能が羽化後最初の1週間とその後の期間で、大きく変容することを示唆している。 ショウジョウバエのキノコ体神経回路を構成する全細胞種の体系的同定Neuronal assemblies of the Drosohila mushroom body.Tanaka, N. K., Tanimoto, H., and Ito, K. J Comp Neurol. (508) 711-755, 2008. (MedLine)
ショウジョウバエ変態期における軸索プルーニングにはアポトーシス細胞貪食遺伝子drprとced-6が必須であるEssential role of the apoptotic cell engulfment genes draper and ced-6 in programmed axon pruning during Drosophila metamorphosis.Takeshi Awasaki, Ryoko Tatsumi, Kuniaki Takahashi, Kunizo Arai, Yoshinobu Nakanishi, Ryu Ueda and Kei Ito.. Neuron (50) 855-867, 2006. (MedLine) 発生過程においておおまかに作られた神経回路を、感覚情報の処理や複雑な行動の制御に最適で、機能的な形に完成させ、維持してゆくためには、いったん作られた神経回路の一部を部分的に作り替えてゆく作業が必要である。その際には、神経細胞の中で不要になった一部の神経線維だけを選択的に取り除いたり、新たな神経線維を再伸長させたりする必要がある。不要な神経線維が特異的に除去されるという現象は、神経回路が形成されるときだけではなく、怪我や病気で神経線維の一部が障害を受けたときにも起きることが知られているが、この現象の仕組みについてはほとんど分かっていない。
ショウジョウバエの脳における聴感覚神経投射の包括的な分類Comprehensive Classification of the Auditory Sensory Projections in the Brain of the Fruit Fly Drosophila melanogaster上川内あづさ、島田尚、伊藤啓 Azusa Kamikouchi, Takashi Shimada and Kei Ito (2006) Journal of Comparative Neurololgy (499) 317-356, 2006. (MedLine)
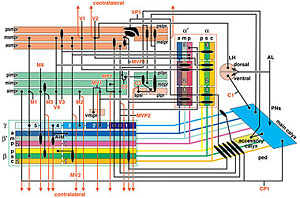
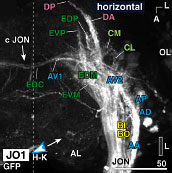
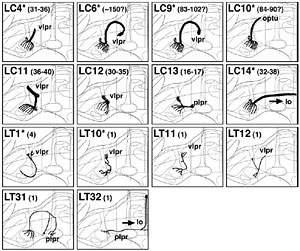
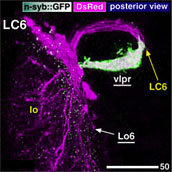
約4,000系統のGAL4エンハンサートラップ系統をスクリーニングして聴覚神経をラベルする系統群を選出し、まずジョンストン器官内部における聴覚神経細胞体の分布を三次元的に解析した。その結果、ジョンストン器官は約500個の聴覚神経を持ち、それらの細胞体は触角第三節の先端部(音受容時の聴覚器官振動の支点)を頂点としてお椀状に分布することを発見した。次にこれら細胞体から脳への軸索投射を解析した結果、軸索投射領域(一次聴覚中枢)は脳本体の中で触角機械感覚野と呼ばれる領域のほぼ全域、vlprと呼ばれる前大脳領域の一部、および食道下神経節の一部にわたって分布している事が分かった。
各ゾーンに投射する聴覚神経を選択的にラベルするGAL4エンハンサートラップ系統群を比較して、ジョンストン器官における細胞体の位置と一次中枢における投射ゾーンとの対応関係を調べたところ、聴覚神経の細胞体は、行き先である各ゾーンに応じて聴覚器官内部のそれぞれ特定の位置に一対のクラスター状、または同心円状に配置していることがわかった。 全40ページ、図版15枚18ページにわたるこの論文によって、これまで明確でなかったショウジョウバエの聴覚一次中枢の領域範囲とその内部構造が、初めて明らかにされた。また、ジョンストン器官の内部に複雑に配置された聴覚細胞の分布パターンが、初めて三次元的に解明された。また、一次聴覚中枢が5つのゾーンに区分けされ、それぞれが感覚器官の特定の領域に分布する特定の神経細胞群からの情報のみを受け取っていることが分かり、これらの領域がそれぞれ異なる種類の聴覚情報処理に関わっている可能性が示唆された。 ショウジョウバエ低次視覚中枢と脳の高次中枢を結ぶ視覚投射神経の体系的解析1:lobula 特異的経路Systematic Analysis of the Visual projection neurons of Drosophila melanogaster - I: Lobula-specific pathways.大綱英生、伊藤啓 Hideo Otsuna and Kei Ito. (2006). Journal of Comparative Neurololgy (497) 928-958. (MedLine) 視覚情報は眼に配置された多数の視細胞から、空間的配置を保ったまま低次視覚中枢に伝えられ、そこで輪郭や動きの抽出など様々な処理が行われる。これらの情報はさらに脳の様々な高次中枢に伝えられ、他の感覚情報と統合される。シンプルな繰り返し構造から成る低次視覚中枢は回路構造の解析が比較的容易であり、これまで詳しく解析されてきた。 神経経路を解析するには、投射先の領域を厳密に区別するための詳細な「住所」が定義されていることが不可欠である。しかし脳が小さく、神経が複雑に錯綜しているショウジョウバエの脳では、キノコ体などごく一部の部分を除き、脳の細かな領域がこれまで厳密に定義されていなかった。そこで我々は、グリア細胞による仕切り構造と容易に同定可能なランドマークになる脳構造を指標にして、脳本体を16の小領域に分割するマップ法をまず定義した。
これらの経路は全部で約500個の細胞から構成される。そのうち8経路約490細胞は、低次視覚中枢において視野の狭い範囲に相当する部分のみにコラム状に投射する神経が数十~百数十個ずつ束になった構造をしており、残り6経路9細胞は、視野全体に接線状に投射する神経が1~4個ずつ存在する構造を取っていた。ラベルされた神経においてシナプス小胞の局在を特異的に可視化して、その分布を解析した結果、前者のコラム状神経はすべて、 31ページ、図版17枚にわたる本論文は、脳内の細かな神経回路をひとつひとつ同定して詳細に記載するという非常に地味な報告であるが、視覚情報が脳でどのように処理されるかを本質的に理解するためには、このような作業を積み重ねて複雑な回路構造の全貌を理解してゆく作業が不可欠である。 ショウジョウバエ変態期における軸索分岐の再編成にはグリア細胞による貪食作用が不可欠であるEngulfing Action of Glial Cells Is Required for Programmed Axon Pruning during Drosophila Metamorphosis粟崎健、伊藤啓 Takeshi Awasaki and Kei Ito. (2004). Current Biology (14) 668-677 (MedLine) 機能的な神経回路の構築には、発生および個体の成熟の過程で神経連絡を局所的に再編成させることが必要である。不要な軸索分岐の消失(プルーニング)はこの過程に見られる特徴的な現象だが、その制御機構はほとんどわかっていない。そこで我々は、変態期に神経回路の大規模な再編成を起こすショウジョウバエの幼虫キノコ体に注目して、軸索分岐消失の過程とそれを制御する機構を解析した。
軸索分岐消失は脱皮ホルモン・エクダイソンの受容で引き起こされることが知られている。そこでエクダイソンの受容をキノコ体神経細胞だけで特異的に阻害したところ、軸索分岐の消失だけでなく、グリア細胞の浸潤も抑制された。つまりグリア細胞の活性化は、グリア自身でなく神経細胞のエクダイソン受容によって制御されている。
ショウジョウバエ2次嗅覚中枢における情報経路の統合Integration of chemosensory pathways in the Drosophila second-order olfactory centers.田中暢明、粟崎健、島田尚、伊藤啓 Nobuaki Karl Tanaka, Takeshi Awasaki, Takashi Shimada, and Kei Ito. (2004). Current Biology (14) 449-457 (MedLine)
そこで我々は、嗅覚情報処理系のモデル系としてショウジョウバエの2次嗅覚中枢における2次神経と3次神経の結合パターンを解析した。まず、昆虫脳の2つの2次嗅覚中枢である側角とキノコ体のうち、側角に注目した。1次中枢である触角葉と2次中枢である側角を結ぶ個々の投射神経(2次神経)が、それぞれの中のどの部分と神経連絡するのかを断面を比較して調べた結果、触角葉の特定の領域から伸びる神経は、側角でも特定の領域に収斂して投射していた。これにより、側角には特定の嗅覚感覚細胞群の情報を選択的に受容するような、いくつかのゾーンが存在することが分かった。
一方、もうひとつの2次中枢であるキノコ体では、2次神経は側角同様にゾーンを作って投射している。しかし3次神経の多くはゾーンをまたがって分岐を広げており、異なるゾーンから情報を集めた多様な3次神経が、キノコ体の同一の領域へと収斂して投射していた。従ってキノコ体経由で嗅覚情報を受ける脳領域は、嗅細胞が受容した全ての種類の嗅覚情報を統合しうるような神経回路構造になっている。
行動実験による脳機能解析から、キノコ体経由の処理経路は匂いの学習や記憶に重要で、一方定型的な匂いへの反応には、側角経由の経路だけで十分なことが分かっている。私たちのデータは、定型的な嗅覚情報処理では匂い情報の全体的統合は行なわれていないことを示唆している。 キノコ体はショウジョウバエの求愛行動に不要であるMushroom bodies are not required for courtship behavior by normal and sexually mosaic Drosophila.木戸麻実、伊藤啓 Asami Kido and Kei Ito. (2002). J Neurobiol (52), 302-11. (MedLine)
しかしよく考えると、これら脳の一部をメス化したハエでは、変化したのは求愛の対象になる相手の性だけであり、複雑な求愛行動のパターン自体は全く変化していない。本当に求愛行動の制御部位をメス化したのならば、オス特異的な行動自体が消失するはずなので、上記の実験結果の解釈には矛盾がある。 そこで我々は、従来のようにたかだか数十の GAL4 系統のスクリーニングではなく、数百系統を大規模にスクリーニングすることによって、transformer を異所発現させたときにオスが求愛行動を行なわなくなる系統を探した。これによって2系統が見つかった。これらの系統でどの細胞がメス化されるのかを知るために、脳内の GAL4 発現パターンを解析した結果、これらの系統ではキノコ体のような特定の領域でなく、脳の非常に広範囲な領域で GAL4 の発現が見られることが分かった。
そこでさらに、「キノコ体での異所発現によってバイセクシャルな行動を引き起こした」という従来の研究で用いられたのと同じ GAL4 系統を用いて、同様にキノコ体の除去を行なった。しかしキノコ体を除去しても、バイセクシャルな行動に変化は生じなかった。またこれらの系統の GAL4 発現パターンを詳しく解析したところ、実際にはキノコ体以外の脳領域でも様々な神経細胞に GAL4 が発現していた。 以上から、従来の通説と異なり、キノコ体はオスの求愛行動の制御に大きな関与をしていないと考えるべきである。求愛行動には嗅覚情報が不可欠であるが、昆虫の嗅覚情報はキノコ体と側角の2つの経路を通じて脳で処理され、そのうち今回の実験ではキノコ体経由の情報伝達を遮断しても影響がないことが分かった。従って求愛行動の制御には、側角経由の情報経路から伝えられる嗅覚情報が十分である可能性が高い。 その他進行中の研究脳の構造脳の発生 脳の機能 脳のインフォマティクス |





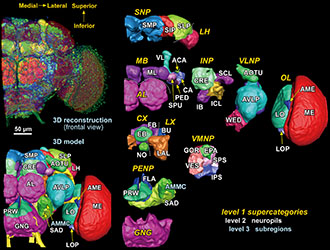 「21 世紀は脳の時代」と言われるように、私たち人間を含む脳の全容を解明する研究が世界各地で盛んに行われ、米国オバマ大統領も昨年宣言した、脳の全ての神経回路を解明する「コネクトーム」と呼ばれる研究が進められている。一方で、脳の大まかな場所の名前は確立しているものの、細かい構造の名前や境界の定義は、研究者や動物種によってばらつきがある。また、研究が盛んな脳の部位では細かい領域分けが整備されているのに対し、あまり盛んでない部位では領域分けや境界の定義が不十分なことがある。このため、脳のどの部分をどう呼ぶかを隅々まで統一した共通基盤が、今後の脳研究を加速させるために求められていた。
「21 世紀は脳の時代」と言われるように、私たち人間を含む脳の全容を解明する研究が世界各地で盛んに行われ、米国オバマ大統領も昨年宣言した、脳の全ての神経回路を解明する「コネクトーム」と呼ばれる研究が進められている。一方で、脳の大まかな場所の名前は確立しているものの、細かい構造の名前や境界の定義は、研究者や動物種によってばらつきがある。また、研究が盛んな脳の部位では細かい領域分けが整備されているのに対し、あまり盛んでない部位では領域分けや境界の定義が不十分なことがある。このため、脳のどの部分をどう呼ぶかを隅々まで統一した共通基盤が、今後の脳研究を加速させるために求められていた。
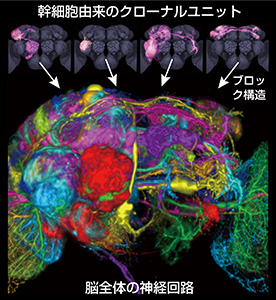 脳にある無数の神経細胞は、全て神経幹細胞が分裂を繰り返して作られる。産み出された神経は、「どの幹細胞から産まれたかという出自に関係なく、それぞれが独自に分化してさまざまな神経回路を作る」と「出自ごとに決まった神経回路を作る」という2つの可能性があるが、実際の脳でどうなっているかは不明だった。そこで神経幹細胞の1つとそれが作る子孫細胞を染め出す実験を繰り返した結果、約100個ある神経幹細胞から作られる子孫細胞群のほとんどが、出自ごとに脳内の決まった場所だけに神経突起の枝を伸ばす特徴的な「クローナルユニット」を形成することが分かった。キノコ体や中心複合体といった特徴的な脳構造や、脳の離れた場所を結ぶ神経線維の束は、特定のクローナルユニットが組み合わさって作られていた。脳全体の神経回路は、このユニットがブロックのように組み合わさって作られている。今回の研究で、ショウジョウバエ脳の全ての場所について、どのユニットがどこに投射し、どのような神経回路構造を作っているかを解明し、複雑な脳構造を持つ生物の神経回路の全体構造を始めて明らかにした。
脳にある無数の神経細胞は、全て神経幹細胞が分裂を繰り返して作られる。産み出された神経は、「どの幹細胞から産まれたかという出自に関係なく、それぞれが独自に分化してさまざまな神経回路を作る」と「出自ごとに決まった神経回路を作る」という2つの可能性があるが、実際の脳でどうなっているかは不明だった。そこで神経幹細胞の1つとそれが作る子孫細胞を染め出す実験を繰り返した結果、約100個ある神経幹細胞から作られる子孫細胞群のほとんどが、出自ごとに脳内の決まった場所だけに神経突起の枝を伸ばす特徴的な「クローナルユニット」を形成することが分かった。キノコ体や中心複合体といった特徴的な脳構造や、脳の離れた場所を結ぶ神経線維の束は、特定のクローナルユニットが組み合わさって作られていた。脳全体の神経回路は、このユニットがブロックのように組み合わさって作られている。今回の研究で、ショウジョウバエ脳の全ての場所について、どのユニットがどこに投射し、どのような神経回路構造を作っているかを解明し、複雑な脳構造を持つ生物の神経回路の全体構造を始めて明らかにした。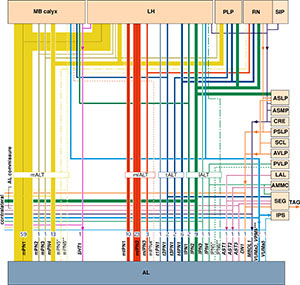 昆虫の嗅覚一次中枢である触角葉は哺乳類の嗅覚一次中枢である嗅球と構造の類似性が非常に高く、その構造・発生・機能は長年にわたって研究されてきた。しかし研究の対象はよく知られた一部の神経種に限られており、嗅覚一次中枢の全神経種を網羅的に同定・解析した研究はこれまでなかった。64ページに及ぶこの論文では、新規に同定した13種類を含む29種類の神経について、一次中枢内部の樹状分岐パターンや二次中枢への投射パターンを網羅的に解明した。特に二次中枢への投射神経については、これまで知られていた3種類の神経投射束と2種類の投射標的領域に加え、新たに4種類の投射神経束と14種類の投射標的領域を発見した。また高次領域の活動情報を一次中枢に伝えるフィードバック神経の候補も、5種類発見した。1つの投射標的領域には複数の神経投射束を介して情報が伝わることがあるが、これら異なる経路の情報が伝わる場所の相関関係も詳しく解析した。一部の神経種に限局されていた従来の研究に対し、神経構造の全貌を網羅した今回の研究は、嗅覚情報処理の様式を理解するのに欠かせない今後の基盤資料となる。
昆虫の嗅覚一次中枢である触角葉は哺乳類の嗅覚一次中枢である嗅球と構造の類似性が非常に高く、その構造・発生・機能は長年にわたって研究されてきた。しかし研究の対象はよく知られた一部の神経種に限られており、嗅覚一次中枢の全神経種を網羅的に同定・解析した研究はこれまでなかった。64ページに及ぶこの論文では、新規に同定した13種類を含む29種類の神経について、一次中枢内部の樹状分岐パターンや二次中枢への投射パターンを網羅的に解明した。特に二次中枢への投射神経については、これまで知られていた3種類の神経投射束と2種類の投射標的領域に加え、新たに4種類の投射神経束と14種類の投射標的領域を発見した。また高次領域の活動情報を一次中枢に伝えるフィードバック神経の候補も、5種類発見した。1つの投射標的領域には複数の神経投射束を介して情報が伝わることがあるが、これら異なる経路の情報が伝わる場所の相関関係も詳しく解析した。一部の神経種に限局されていた従来の研究に対し、神経構造の全貌を網羅した今回の研究は、嗅覚情報処理の様式を理解するのに欠かせない今後の基盤資料となる。
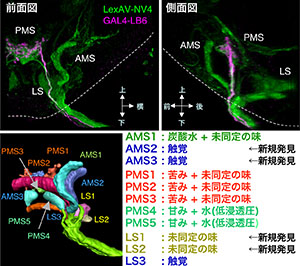 当研究室ではこれまで視覚・嗅覚・聴覚の感覚中枢の詳細な構造を解析してきたが、これに引き続き、残る重要な感覚情報処理機構の1つである味覚中枢の詳細な解析を行った。昆虫では味覚感覚神経の軸索が脳に直接投射するため、原理的には味覚中枢における詳細な感覚マップを作ることが可能である。しかし、視覚中枢や嗅覚中枢がグリア細胞などで仕切られた明瞭な構造を持つのに比べ、味覚中枢は複雑な構造で、かつ周囲との境界が明瞭でないため、これまで味覚中枢の全体構造もきちんとした投射マップも分かっていなかった。そこで本研究では、従来から用いられてきたGAL4に加えて、LexAを用いた発現誘導系を使って新たに多数の系統を作成して、味覚中枢を特異的にラベルする系統を選び出した。特定の味覚神経をラベルするさまざまなGAL4とLexA系統で二重ラベルを行うことによって、初の体系的な味覚中枢の神経投射マップを構築した。その結果、甘みと水の情報は味覚中枢の重複する領域に送られること、甘みと水/苦み/炭酸の3つの情報はそれぞれ異なる領域に送られること、味覚情報と「口が対象物に接触した」ことを検知する触覚情報は、隣接するが異なる領域に送られること、といった味覚情報処理の基本構造が判明した。
当研究室ではこれまで視覚・嗅覚・聴覚の感覚中枢の詳細な構造を解析してきたが、これに引き続き、残る重要な感覚情報処理機構の1つである味覚中枢の詳細な解析を行った。昆虫では味覚感覚神経の軸索が脳に直接投射するため、原理的には味覚中枢における詳細な感覚マップを作ることが可能である。しかし、視覚中枢や嗅覚中枢がグリア細胞などで仕切られた明瞭な構造を持つのに比べ、味覚中枢は複雑な構造で、かつ周囲との境界が明瞭でないため、これまで味覚中枢の全体構造もきちんとした投射マップも分かっていなかった。そこで本研究では、従来から用いられてきたGAL4に加えて、LexAを用いた発現誘導系を使って新たに多数の系統を作成して、味覚中枢を特異的にラベルする系統を選び出した。特定の味覚神経をラベルするさまざまなGAL4とLexA系統で二重ラベルを行うことによって、初の体系的な味覚中枢の神経投射マップを構築した。その結果、甘みと水の情報は味覚中枢の重複する領域に送られること、甘みと水/苦み/炭酸の3つの情報はそれぞれ異なる領域に送られること、味覚情報と「口が対象物に接触した」ことを検知する触覚情報は、隣接するが異なる領域に送られること、といった味覚情報処理の基本構造が判明した。
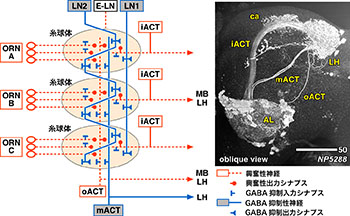 GABAを伝達物質とする抑制性神経は神経回路の演算機能に重要な役割を果たす。抑制性情報の送信と受信が神経回路のどの部分で行われるのかを詳しく解析するため、我々はGABA伝達物質を放出する神経とGABA信号を受容する入力シナプスを持つ神経の分布を、ショウジョウバエ脳において詳細に解析した。GABAを放出する神経の数は脳の数カ所にクラスターを作っていたのに対し、GABA信号を受容する入力シナプスはほとんどの細胞に存在した。特に、神経回路の構成がよく分かっている嗅覚一次中枢において蛍光in situハイブリダイゼーション法と特定の神経種でのGFP発現誘導の二重染色解析を行った結果、形態が異なる2種類の局所神経と、1種類の投射神経のみが特異的にGABA抑制性であり、このうち投射神経は一次中枢内には出力シナプスを持っていなかった。従って一次中枢での抑制性情報は、局所神経のみによって発信されている。一方GABA受容入力シナプスは全ての神経種に存在し、抑制性信号の送信側と受信側には大きな差があることが分かった。
GABAを伝達物質とする抑制性神経は神経回路の演算機能に重要な役割を果たす。抑制性情報の送信と受信が神経回路のどの部分で行われるのかを詳しく解析するため、我々はGABA伝達物質を放出する神経とGABA信号を受容する入力シナプスを持つ神経の分布を、ショウジョウバエ脳において詳細に解析した。GABAを放出する神経の数は脳の数カ所にクラスターを作っていたのに対し、GABA信号を受容する入力シナプスはほとんどの細胞に存在した。特に、神経回路の構成がよく分かっている嗅覚一次中枢において蛍光in situハイブリダイゼーション法と特定の神経種でのGFP発現誘導の二重染色解析を行った結果、形態が異なる2種類の局所神経と、1種類の投射神経のみが特異的にGABA抑制性であり、このうち投射神経は一次中枢内には出力シナプスを持っていなかった。従って一次中枢での抑制性情報は、局所神経のみによって発信されている。一方GABA受容入力シナプスは全ての神経種に存在し、抑制性信号の送信側と受信側には大きな差があることが分かった。
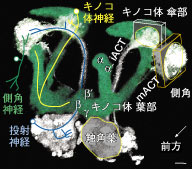
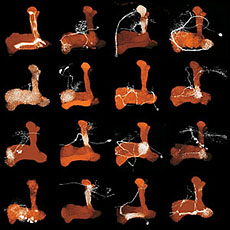 ショウジョウバエのキノコ体は学習・記憶の研究に広く利用されているモデル神経系で、キノコ体を形成する主要神経であるKenyon神経の構造や機能については、以前から詳しい解析が行われている。しかしKenyon神経だけでは、キノコ体は外部と何の情報のやりとりもできない。それで我々は6年をかけて、Kenyon神経だけでなくそれ以外の内部神経、さらにキノコ体の内部と外部を接続する神経を含んだキノコ体の全ての神経細胞種について、体系的な同定と投射マップの作成を行った。
まずKenyon神経の様々な種類を同定解析した結果、先端のlobe部分はα’/β’ lobe, α/β lobeがそれぞれ細胞種ごとに3層に分かれているのに対し、γ lobeは一様な構造だった。これら7種の神経のうち、6種は基部にあるcalyx部の同じ領域に重複して樹状突起を伸ばしていた。この領域は嗅覚一次中枢から濃密な投射が見られるので、これら6種の神経はほぼ同一内容の嗅覚情報を重複して受け取っている。一方、α/β lobeの最外層の神経だけは、calyxから突き出た特定の領域のみに樹状突起を伸ばしていた。この領域には嗅覚神経は一切投射しないため、このタイプのKenyon細胞は独自の機能を持っていると推定される。また、キノコ体の内部のみに投射するKenyon神経以外の神経を、既知の1種を含む3種類同定したが、これらはいずれもKenyon神経と異なりlobeのすべての層にまたがって投射して、Kenyon神経が形成する層の間を結んでいた。
ショウジョウバエのキノコ体は学習・記憶の研究に広く利用されているモデル神経系で、キノコ体を形成する主要神経であるKenyon神経の構造や機能については、以前から詳しい解析が行われている。しかしKenyon神経だけでは、キノコ体は外部と何の情報のやりとりもできない。それで我々は6年をかけて、Kenyon神経だけでなくそれ以外の内部神経、さらにキノコ体の内部と外部を接続する神経を含んだキノコ体の全ての神経細胞種について、体系的な同定と投射マップの作成を行った。
まずKenyon神経の様々な種類を同定解析した結果、先端のlobe部分はα’/β’ lobe, α/β lobeがそれぞれ細胞種ごとに3層に分かれているのに対し、γ lobeは一様な構造だった。これら7種の神経のうち、6種は基部にあるcalyx部の同じ領域に重複して樹状突起を伸ばしていた。この領域は嗅覚一次中枢から濃密な投射が見られるので、これら6種の神経はほぼ同一内容の嗅覚情報を重複して受け取っている。一方、α/β lobeの最外層の神経だけは、calyxから突き出た特定の領域のみに樹状突起を伸ばしていた。この領域には嗅覚神経は一切投射しないため、このタイプのKenyon細胞は独自の機能を持っていると推定される。また、キノコ体の内部のみに投射するKenyon神経以外の神経を、既知の1種を含む3種類同定したが、これらはいずれもKenyon神経と異なりlobeのすべての層にまたがって投射して、Kenyon神経が形成する層の間を結んでいた。
 さらに、キノコ体を周囲の他の脳領域とを結ぶ外部神経を新たに多数同定して、それらの構造を解析した。キノコ体は嗅覚とそれ以外の感覚情報の連合学習に重要だとされているが、嗅覚情報は一次中枢から直接入射しているのに対し、視覚・聴覚・味覚などの感覚中枢と直接連絡するような外部神経はひとつも見られなかった。また、嗅覚投射神経やKenyon神経は左右半球が独立しているのに対し、外部神経は左右の相同なキノコ体領域を結ぶものが過半数であった。Kenyon神経がlobeの長軸方向を貫通するように投射するのに対し、これら外部神経は全てlobeを長軸方向に2~5分割するセグメント状の範囲のみに投射していた。この結果、キノコ体には層状に投射するKenyon神経とセグメント状に投射する外部神経によって、多数のマトリクス状の小部位が形成されており、1つのKenyon細胞は長軸方向に複数の種類の外部神経と相互作用することが分かった。論文というよりも図鑑に近いこの仕事によって、これまで曖昧だったキノコ体内部の神経回路構造を、初めて詳しく明らかにすることができた。
さらに、キノコ体を周囲の他の脳領域とを結ぶ外部神経を新たに多数同定して、それらの構造を解析した。キノコ体は嗅覚とそれ以外の感覚情報の連合学習に重要だとされているが、嗅覚情報は一次中枢から直接入射しているのに対し、視覚・聴覚・味覚などの感覚中枢と直接連絡するような外部神経はひとつも見られなかった。また、嗅覚投射神経やKenyon神経は左右半球が独立しているのに対し、外部神経は左右の相同なキノコ体領域を結ぶものが過半数であった。Kenyon神経がlobeの長軸方向を貫通するように投射するのに対し、これら外部神経は全てlobeを長軸方向に2~5分割するセグメント状の範囲のみに投射していた。この結果、キノコ体には層状に投射するKenyon神経とセグメント状に投射する外部神経によって、多数のマトリクス状の小部位が形成されており、1つのKenyon細胞は長軸方向に複数の種類の外部神経と相互作用することが分かった。論文というよりも図鑑に近いこの仕事によって、これまで曖昧だったキノコ体内部の神経回路構造を、初めて詳しく明らかにすることができた。
 ショウジョウバエでは、蛹(サナギ)の時期に脳内の神経回路が幼虫型から成虫型に作り替えられる。匂いに関する学習に中心的な役割を果たしている「キノコ体」と呼ばれる脳領域では、幼虫期に作られた神経回路の一部が蛹の初期に特異的に除去され、その後成虫型の神経回路が新たに形成されることが知られている。これまでの研究で、これらの神経線維が的確なタイミングで正確に除去されるためには、神経の自発的変性だけでなく、周囲のグリア細胞がこれらの神経線維を選択的に取り囲んで消化する「貪食作用」が必要であることが明らかになった(本ページの下方参照)。その後、脊椎動物においても、不要な神経線維の選択的な除去において、グリア細胞の貪食作用が関与していることが報告され、グリア細胞による不要な神経線維の除去が、神経回路の形成と維持において普遍的な現象であることが示された。しかし、グリア細胞がどのような分子機構によって不要となった神経細胞を認識して、選択的に食べているのかについては、ほとんど分かっていなかった。
ショウジョウバエでは、蛹(サナギ)の時期に脳内の神経回路が幼虫型から成虫型に作り替えられる。匂いに関する学習に中心的な役割を果たしている「キノコ体」と呼ばれる脳領域では、幼虫期に作られた神経回路の一部が蛹の初期に特異的に除去され、その後成虫型の神経回路が新たに形成されることが知られている。これまでの研究で、これらの神経線維が的確なタイミングで正確に除去されるためには、神経の自発的変性だけでなく、周囲のグリア細胞がこれらの神経線維を選択的に取り囲んで消化する「貪食作用」が必要であることが明らかになった(本ページの下方参照)。その後、脊椎動物においても、不要な神経線維の選択的な除去において、グリア細胞の貪食作用が関与していることが報告され、グリア細胞による不要な神経線維の除去が、神経回路の形成と維持において普遍的な現象であることが示された。しかし、グリア細胞がどのような分子機構によって不要となった神経細胞を認識して、選択的に食べているのかについては、ほとんど分かっていなかった。
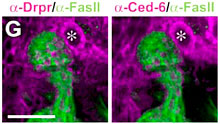
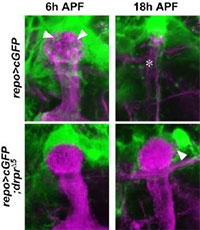 今回の研究で、まず蛹初期のショウジョウバエのグリア細胞が幼虫のキノコ体神経線維を貪食する際に、これらのグリア細胞でdraper (drpr) と ced-6 という2つの遺伝子が一時的に強く発現していることを発見した。drpr遺伝子はスカベンジャー受容体分子(Drprタンパク質)を、ced-6 遺伝子はアダプター分子(Ced-6タンパク質)をそれぞれ作り、これらの遺伝子は、線虫においてプログラム細胞死(アポトーシス)を起こした細胞を、食細胞が貪食する際に必要であることが知られている。RNA干渉法を利用して、これらの遺伝子の機能をグリア細胞のみで特異的に阻害したところ、グリア細胞による貪食作用が抑制され、幼虫のキノコ体神経線維が除去されなくなった。また、RNA干渉法でなくdrpr遺伝子の機能が失われた突然変異系統を使った実験でも、同様の現象が観察された。一方で、グリア細胞によって除去される幼虫キノコ体神経線維では、グリア細胞の貪食作用とは独立に微小管細胞骨格の崩壊という変性現象が起こっており、この現象はキノコ体神経細胞がエクダイソンを受容することで誘導されることも見出した。
今回の研究で、まず蛹初期のショウジョウバエのグリア細胞が幼虫のキノコ体神経線維を貪食する際に、これらのグリア細胞でdraper (drpr) と ced-6 という2つの遺伝子が一時的に強く発現していることを発見した。drpr遺伝子はスカベンジャー受容体分子(Drprタンパク質)を、ced-6 遺伝子はアダプター分子(Ced-6タンパク質)をそれぞれ作り、これらの遺伝子は、線虫においてプログラム細胞死(アポトーシス)を起こした細胞を、食細胞が貪食する際に必要であることが知られている。RNA干渉法を利用して、これらの遺伝子の機能をグリア細胞のみで特異的に阻害したところ、グリア細胞による貪食作用が抑制され、幼虫のキノコ体神経線維が除去されなくなった。また、RNA干渉法でなくdrpr遺伝子の機能が失われた突然変異系統を使った実験でも、同様の現象が観察された。一方で、グリア細胞によって除去される幼虫キノコ体神経線維では、グリア細胞の貪食作用とは独立に微小管細胞骨格の崩壊という変性現象が起こっており、この現象はキノコ体神経細胞がエクダイソンを受容することで誘導されることも見出した。
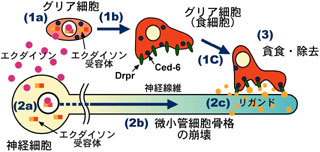 以上の解析結果により、キノコ体神経細胞はエクダイソンを受容し、特定の神経線維の細胞骨格を変性させ、グリア細胞がDrprタンパク質とCed-6タンパク質の働きによって、この変性神経線維を特異的に検出して、貪食・除去しているというメカニズムが示唆される。この研究から、グリア細胞では、プログラム細胞死を起こして死んだ細胞の除去だけでなく、生きている神経細胞の一部である変性神経線維の除去にも、同じ分子機構が使われているという新しい概念が提唱された。
以上の解析結果により、キノコ体神経細胞はエクダイソンを受容し、特定の神経線維の細胞骨格を変性させ、グリア細胞がDrprタンパク質とCed-6タンパク質の働きによって、この変性神経線維を特異的に検出して、貪食・除去しているというメカニズムが示唆される。この研究から、グリア細胞では、プログラム細胞死を起こして死んだ細胞の除去だけでなく、生きている神経細胞の一部である変性神経線維の除去にも、同じ分子機構が使われているという新しい概念が提唱された。
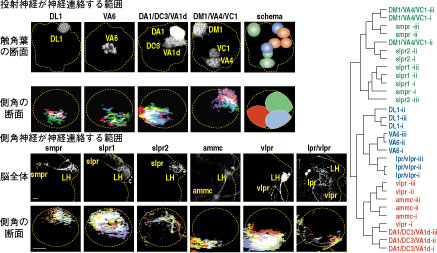
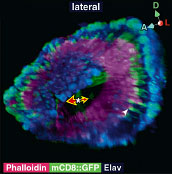 ショウジョウバエは、雄が雌に対して種特異的な羽音を使って求愛することから、音を介した個体間コミュニケーションや脳における聴覚情報処理を研究する上で興味深い材料である。聴覚情報が脳でどのように処理されるかを解明する上で、音情報処理に関わる神経回路構造の同定は必要不可欠である。しかし、昆虫脳において聴覚神経が脳へどのように投射するかを単一細胞レベルで詳しく調べた研究はほとんどない。そこで我々は、触角の第二節に存在する昆虫の聴覚器官(ジョンストン器官)から脳の一次聴覚中枢へ投射する、聴覚神経の回路構造を体系的に解析した。
ショウジョウバエは、雄が雌に対して種特異的な羽音を使って求愛することから、音を介した個体間コミュニケーションや脳における聴覚情報処理を研究する上で興味深い材料である。聴覚情報が脳でどのように処理されるかを解明する上で、音情報処理に関わる神経回路構造の同定は必要不可欠である。しかし、昆虫脳において聴覚神経が脳へどのように投射するかを単一細胞レベルで詳しく調べた研究はほとんどない。そこで我々は、触角の第二節に存在する昆虫の聴覚器官(ジョンストン器官)から脳の一次聴覚中枢へ投射する、聴覚神経の回路構造を体系的に解析した。
 聴覚神経の脳内での投射パターンを解析したところ、一次聴覚中枢の内部は少なくとも5つのゾーンに分かれており、それら5つのゾーンはさらに19個のサブエリアに分割できることを見いだした。軸索の中で出力シナプスが局在する部位だけを可視化したところ、5つのゾーンには全てシナプス出力部位が存在していた。さらに、熱ショックフリップアウト法を用いて個々の聴覚神経の軸索投射を可視化して解析したところ、それぞれの細胞はこれら5つのゾーンのうちの、どれか1つのゾーンのみに投射していた。つまり、個々の聴覚神経は一次聴覚中枢の中の特定のゾーンのみに情報を伝達している。
聴覚神経の脳内での投射パターンを解析したところ、一次聴覚中枢の内部は少なくとも5つのゾーンに分かれており、それら5つのゾーンはさらに19個のサブエリアに分割できることを見いだした。軸索の中で出力シナプスが局在する部位だけを可視化したところ、5つのゾーンには全てシナプス出力部位が存在していた。さらに、熱ショックフリップアウト法を用いて個々の聴覚神経の軸索投射を可視化して解析したところ、それぞれの細胞はこれら5つのゾーンのうちの、どれか1つのゾーンのみに投射していた。つまり、個々の聴覚神経は一次聴覚中枢の中の特定のゾーンのみに情報を伝達している。
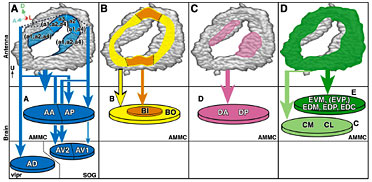 このことにより、視覚、嗅覚、味覚など他の感覚受容器と同様に、聴覚においても感覚器官と脳の一次中枢の間に厳密な相関マップが存在する事が示された。
このことにより、視覚、嗅覚、味覚など他の感覚受容器と同様に、聴覚においても感覚器官と脳の一次中枢の間に厳密な相関マップが存在する事が示された。
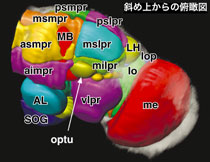 しかし低次視覚中枢と高次中枢を結ぶ神経の回路構造については、いまだ知見が非常に限られている。そこで我々は視覚情報処理のモデル系として、ショウジョウバエの低次視覚中枢である視葉と、高次中枢が集積する脳本体との間を結ぶ視覚投射神経を、体系的に同定解析した。
しかし低次視覚中枢と高次中枢を結ぶ神経の回路構造については、いまだ知見が非常に限られている。そこで我々は視覚情報処理のモデル系として、ショウジョウバエの低次視覚中枢である視葉と、高次中枢が集積する脳本体との間を結ぶ視覚投射神経を、体系的に同定解析した。
 さらに、4,000系統のGAL4エンハンサートラップ系統をスクリーニングして視覚投射神経をラベルする系統を選び、それらがラベルする神経を比較した結果、合計44種類の視覚投射神経を同定した。量が多いので4部に分けて発表することとし、第1部にあたる本論文では、まず視葉の中のlobula領域と脳本体を結ぶ14経路について報告した。
さらに、4,000系統のGAL4エンハンサートラップ系統をスクリーニングして視覚投射神経をラベルする系統を選び、それらがラベルする神経を比較した結果、合計44種類の視覚投射神経を同定した。量が多いので4部に分けて発表することとし、第1部にあたる本論文では、まず視葉の中のlobula領域と脳本体を結ぶ14経路について報告した。
 出力シナプスが高次中枢側にある求心性経路だったのに対し、後者の接線状神経では約半数が、出力シナプスが低次中枢側にある遠心性経路であった。これらは、高次中枢が低次中枢の作動を修飾するための制御経路になっていると推測される。また、前者のほとんどは脳本体の中でvlprと呼ばれる領域に投射していたのに対し、後者はplprから投射しており、その分布は大きく異なっていた。
出力シナプスが高次中枢側にある求心性経路だったのに対し、後者の接線状神経では約半数が、出力シナプスが低次中枢側にある遠心性経路であった。これらは、高次中枢が低次中枢の作動を修飾するための制御経路になっていると推測される。また、前者のほとんどは脳本体の中でvlprと呼ばれる領域に投射していたのに対し、後者はplprから投射しており、その分布は大きく異なっていた。
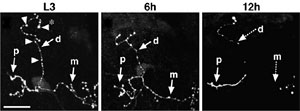 まず幼虫キノコ体の単一細胞のみをラベルして、変態期における軸索の形態変化を詳細に解析した。その結果、軸索上の「バリコシティー」と呼ばれるこぶ状の構造体がまず消失し、次いで軸索の断片化が起こり、軸索分岐全体の消失へと進むことが分かった。これにより、軸索分岐消失は軸索の先端から根本への退縮ではなく、軸索の断裂・変性によって起こることが明らかになった。
まず幼虫キノコ体の単一細胞のみをラベルして、変態期における軸索の形態変化を詳細に解析した。その結果、軸索上の「バリコシティー」と呼ばれるこぶ状の構造体がまず消失し、次いで軸索の断片化が起こり、軸索分岐全体の消失へと進むことが分かった。これにより、軸索分岐消失は軸索の先端から根本への退縮ではなく、軸索の断裂・変性によって起こることが明らかになった。
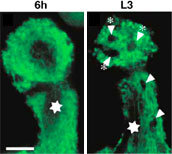 次に単一細胞でなく800個のキノコ体神経の軸索束全体をラベルして解析した結果、隣接する複数の軸索分岐に存在するバリコシティーが、集団でクラスター状に消失することが分かった。これは、軸索分岐消失が軸索1本単位で起こるのではなく、隣り合った異なる軸索の間で同時に起こることを示唆する。そこで我々は、軸索分岐消失が何らかの外部要因によって引き起こされている可能性を考え、キノコ体周囲のグリア細胞群に注目した。変態期における形態変化を調べたところ、幼虫期には軸索束の外周を覆っていたグリア細胞は、変態期に入ると軸索束の間に細胞突起を浸潤させ、バリコシティーをクラスター状に包み込み、貪食して分解していた。
次に単一細胞でなく800個のキノコ体神経の軸索束全体をラベルして解析した結果、隣接する複数の軸索分岐に存在するバリコシティーが、集団でクラスター状に消失することが分かった。これは、軸索分岐消失が軸索1本単位で起こるのではなく、隣り合った異なる軸索の間で同時に起こることを示唆する。そこで我々は、軸索分岐消失が何らかの外部要因によって引き起こされている可能性を考え、キノコ体周囲のグリア細胞群に注目した。変態期における形態変化を調べたところ、幼虫期には軸索束の外周を覆っていたグリア細胞は、変態期に入ると軸索束の間に細胞突起を浸潤させ、バリコシティーをクラスター状に包み込み、貪食して分解していた。
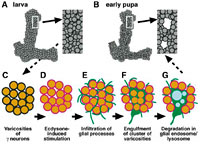 このグリア細胞の活動が軸索分岐消失に必須なのかを調べるため、遺伝子異所発現によって貪食などの細胞機能をグリア特異的に阻害してみた。すると、単にグリア細胞の軸索束への浸潤が抑制されただけでなく、神経の軸索分岐消失も著しく阻害された。従って、グリア細胞は単に神経細胞が自律的に断裂変性させた軸索分岐の残骸を処理しているのではなく、積極的に軸索分岐を貪食・分解していることが分かった。
このグリア細胞の活動が軸索分岐消失に必須なのかを調べるため、遺伝子異所発現によって貪食などの細胞機能をグリア特異的に阻害してみた。すると、単にグリア細胞の軸索束への浸潤が抑制されただけでなく、神経の軸索分岐消失も著しく阻害された。従って、グリア細胞は単に神経細胞が自律的に断裂変性させた軸索分岐の残骸を処理しているのではなく、積極的に軸索分岐を貪食・分解していることが分かった。
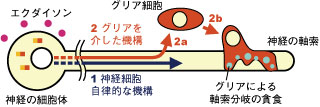 軸索分岐消失に関する従来の研究は、“神経細胞の細胞自律的な制御機構(1)”だけに着目してきた。しかしキノコ体神経における我々の研究は、 (2a)神経細胞がグリア細胞を活性化し、(2b)グリア細胞が軸策束に浸潤して積極的に軸索分岐を貪食・分解する、という“神経-グリア相互作用による制御機構”が非常に重要であることを提唱した。
軸索分岐消失に関する従来の研究は、“神経細胞の細胞自律的な制御機構(1)”だけに着目してきた。しかしキノコ体神経における我々の研究は、 (2a)神経細胞がグリア細胞を活性化し、(2b)グリア細胞が軸策束に浸潤して積極的に軸索分岐を貪食・分解する、という“神経-グリア相互作用による制御機構”が非常に重要であることを提唱した。
 生物が匂いの種類を認識し、適切な行動を取るためには、嗅覚感覚器にある数十~数千種類の感覚細胞からの情報を高次中枢で統合する必要がある。最近の研究で、感覚細胞(1次神経)は種類ごとに1次嗅覚中枢のなかの特定の領域(糸球体)に投射し、さらに2次神経も1次嗅覚中枢から2次中枢へ、決まった様式で投射することが分かってきた。しかし、2次神経によって伝えられた匂い特異的な信号を、さらに高次の神経がどのように読みとっているのかについては、まったく分かっていない。
生物が匂いの種類を認識し、適切な行動を取るためには、嗅覚感覚器にある数十~数千種類の感覚細胞からの情報を高次中枢で統合する必要がある。最近の研究で、感覚細胞(1次神経)は種類ごとに1次嗅覚中枢のなかの特定の領域(糸球体)に投射し、さらに2次神経も1次嗅覚中枢から2次中枢へ、決まった様式で投射することが分かってきた。しかし、2次神経によって伝えられた匂い特異的な信号を、さらに高次の神経がどのように読みとっているのかについては、まったく分かっていない。
 さらに、従来ほとんど知られていなかった3次神経の構造を解析した。側角では、3次神経は上記のゾーンのどれかひとつの範囲内に分岐を広げており、それぞれのゾーンに由来する3次神経は、脳の別々の領域へと投射していた。上手の右端のようにクラスター解析を行なうと、触角葉と高次脳の特異的な領域同士が、深く相関していることが分かった。従って、側角経由で嗅覚情報を受けるさらに高次の脳領域は、ひとつのゾーンに伝えられる限られた種類の嗅覚情報しか統合できないという神経回路構造になっている。
さらに、従来ほとんど知られていなかった3次神経の構造を解析した。側角では、3次神経は上記のゾーンのどれかひとつの範囲内に分岐を広げており、それぞれのゾーンに由来する3次神経は、脳の別々の領域へと投射していた。上手の右端のようにクラスター解析を行なうと、触角葉と高次脳の特異的な領域同士が、深く相関していることが分かった。従って、側角経由で嗅覚情報を受けるさらに高次の脳領域は、ひとつのゾーンに伝えられる限られた種類の嗅覚情報しか統合できないという神経回路構造になっている。
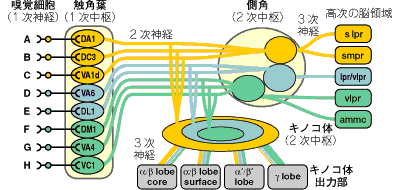 本研究は3次神経の神経回路構造を解明したはじめての研究であり、機能が異なる2つの2次嗅覚中枢で、3次神経によって統合される嗅覚情報の範囲に大きな違いがあることを明らかにした。特定の匂い分子群を受容する嗅細胞からの神経は、触角葉の特定の領域にまとめて入射する。そこから伸びる投射神経は、上の図に示すように側角の特定の領域に伸び、さらに特定の高次脳領域へと送られてゆく。嗅細胞全体からの情報は統合されず、いくつかの流れに分かれたままである。一方キノコ体では、投射神経は特定の領域に入射するものの、それを受けるキノコ体神経は幅広い領域からの情報を4種類のキノコ体出力部にまとめて伝えるので、出力部では嗅細胞全体からの情報が統合されている。
本研究は3次神経の神経回路構造を解明したはじめての研究であり、機能が異なる2つの2次嗅覚中枢で、3次神経によって統合される嗅覚情報の範囲に大きな違いがあることを明らかにした。特定の匂い分子群を受容する嗅細胞からの神経は、触角葉の特定の領域にまとめて入射する。そこから伸びる投射神経は、上の図に示すように側角の特定の領域に伸び、さらに特定の高次脳領域へと送られてゆく。嗅細胞全体からの情報は統合されず、いくつかの流れに分かれたままである。一方キノコ体では、投射神経は特定の領域に入射するものの、それを受けるキノコ体神経は幅広い領域からの情報を4種類のキノコ体出力部にまとめて伝えるので、出力部では嗅細胞全体からの情報が統合されている。
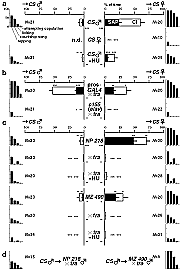 ショウジョウバエでは、オスがメスに対し積極的かつ定型的な求愛行動を行う。このような雌雄で明確に異なる行動は、脳の中の雌雄で異なる構造や特性を持つ領域によって制御されている可能性があり、脳の構造と機能の相関を調べるモデル系として関心を集めてきた。従来の研究で、昆虫の性決定遺伝子である transformer 遺伝子を GAL4 系統を用いてキノコ体など脳の一部で強制発現させてオスの脳の一部をメス化したところ、オスがメスだけでなくオスも求愛するバイセクシャルな行動を取るようになったことから、キノコ体がハエの求愛行動を制御する座として重要であるという仮説が強く提唱されていた。
ショウジョウバエでは、オスがメスに対し積極的かつ定型的な求愛行動を行う。このような雌雄で明確に異なる行動は、脳の中の雌雄で異なる構造や特性を持つ領域によって制御されている可能性があり、脳の構造と機能の相関を調べるモデル系として関心を集めてきた。従来の研究で、昆虫の性決定遺伝子である transformer 遺伝子を GAL4 系統を用いてキノコ体など脳の一部で強制発現させてオスの脳の一部をメス化したところ、オスがメスだけでなくオスも求愛するバイセクシャルな行動を取るようになったことから、キノコ体がハエの求愛行動を制御する座として重要であるという仮説が強く提唱されていた。
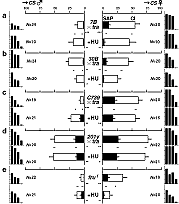 ショウジョウバエでは、発生過程の特定の時期に分裂阻害剤であるヒドロキシウレア(HU)を投与することにより、キノコ体を形成する神経幹細胞とその子孫細胞だけを、特異的に除去することができる。transformer 遺伝子の異所発現によって求愛行動が消失する上記の2系統は、この方法でキノコ体を除去しても、行動パターンに変化は見られなかった。また、野生型(CS 系統)のハエのキノコ体を除去しても、メスにしか求愛しないという行動パターンに変化は見られなかった。
ショウジョウバエでは、発生過程の特定の時期に分裂阻害剤であるヒドロキシウレア(HU)を投与することにより、キノコ体を形成する神経幹細胞とその子孫細胞だけを、特異的に除去することができる。transformer 遺伝子の異所発現によって求愛行動が消失する上記の2系統は、この方法でキノコ体を除去しても、行動パターンに変化は見られなかった。また、野生型(CS 系統)のハエのキノコ体を除去しても、メスにしか求愛しないという行動パターンに変化は見られなかった。