|
|
|
|
|

|
遺伝学の代表的な実験動物として100年間使われてきたという研究の蓄積を持つショウジョウバエでは、掛け合わせによって様々な遺伝子を合わせ持った個体を作る作業が、非常に簡単に行なえます。GFP などの遺伝子を組み込んで細胞をラベルする実験は多くの動物で行なわれていますが、それを発展させてここに示すような多彩な細胞ラベルが可能なのは、現在のところキイロショウジョウバエだけです。 応用編1:レポーター遺伝子の使い分けによる、異なった細胞構造の可視化遺伝子発現を用いた通常の細胞ラベル法では、「1:どの細胞で 2:どの遺伝子を」発現させるかが一体化してしまっています。ある遺伝子を特異的に発現する系統があったとき、その同じ細胞で別の遺伝子を発現させようと思っても、それには発現ベクターの DNA コンストラクトの作り直しや、新たな形質転換系統樹立の作業など、膨大な実験の手間が必要です。 GAL4 エンハンサートラップ系統では、「どの細胞で発現させるか」と「どの遺伝子を発現させるか」が分かれています。GAL4 転写調節因子の標的配列である UAS の後ろに、どんな遺伝子を組み合わせるかによって、自由に発現させる遺伝子を選ぶことができます。 1980年代から90年代初頭にかけてレポーターとして広く用いられた lacZ 遺伝子は、産物であるβガラクトシダーゼの分子量が大きく、しかも4量体になって働くため、細胞質内での拡散が悪く、細い線維の先までラベルできないという欠点がありました。そこで、軸索輸送で線維の先端まで運ばれる細胞骨格タンパクであるタウやキネシンの遺伝子に lacZ を融合させて動的に輸送させる方法が開発されました (UAS-tau::lacZ, UAS-kinesin::lacZ)。しかし巨大なβガラクトシダーゼを無理やり細胞内で移動させることは細胞へのダメージが大きく、レポーター遺伝子の発現によって細胞が死んでしまったり、形態に異常が起きたりする欠点が見つかってきました。
GFP の登場1990年代に登場した、自ら蛍光を発するクラゲの緑色蛍光色素 GFP は、出現当時多くの注目を集めました。しかし当初のものは非常に暗く、また蛍光観察するとすぐに褪色してしまい、ほとんど実用になりませんでした。多くの研究者が改良を重ねた結果、活性部位のアミノ酸を人工置換して蛍光強度と抗褪色性を向上させた改良版が開発され、普及したのが、1996年ごろのことです。GFP は分子量が小さく、しかも単量体で働くので、非常に良好に細胞内に拡散します。そのため、神経線維の先もきれいに染まります。発現によって細胞に与えるダメージも非常に小さいのが特長です。 GFP は単体でも蛍光を発し、可視化できますが、蛍光標本でなく明視野やノマルスキー検鏡の標本を作りたい場合や二重染色を行ないたい場合には、抗 GFP 抗体でラベルすることができると便利です。当初は GFP をラベルする良い抗体がなく、まともな抗体染色は不可能でした。しかし2000年ごろから良い抗体が市販されるようになり、これによって GFP は、観察法を問わずオールラウンドなレポーターとしての地位を確立しました。 さらに細胞末端部をきれいにラベルするために、膜タンパク CD8 と融合させて細胞膜表面だけをラベルする手法も登場しました (UAS-mCF8::GFP)。これは細胞質でなく膜だけをラベルするので、細胞体積のわりに細胞膜の表面積が少ない細胞本体や太い軸索は比較的薄くラベルされ、逆に体積のわりに細胞膜の表面積が多い、細い繊維の末端部は、濃いめにラベルされるという特徴があります。通常の GFP が神経線維の「幹」を強く、「枝」を弱く染めるのに対し、mCD8::GFP は、「枝」を強く、「幹」を弱く染めるわけです。また、シナプスなどで形成された枝の上の「コブ」構造は、GFP ではよく見えますが mCD8::GFP ではよく見えません。
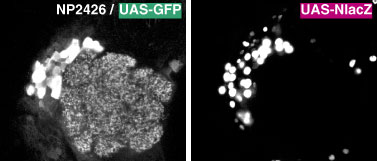
細胞が密集している領域では、細胞全体をラベルすると隣り合う細胞のラベルが癒合してしまいます。細胞の本体だけをラベルするために、ウィルスの核移行タンパク遺伝子を融合させて核だけをラベルする方法も開発されました。GFP とつなげた UAS-nuclear GFP と、lacZ とつなげた UAS-NlacZ とがあります。nuclear GFP は何らかの原因で細胞核以外にもラベルが漏れてしまうのに対し、NlacZ はラベルが真に核に局在するので、後者の方が安定して利用できるレポーターです。
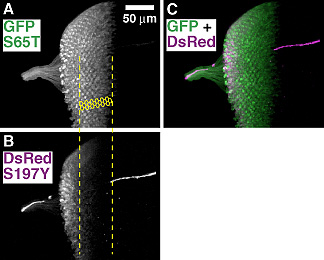
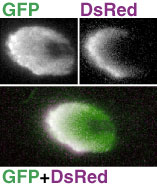
単に細胞の形態でなく、機能部位に応じて染め分ける方法もあります。本研究室ではアメリカのグループと共同で、シナプス小胞がシナプス前膜にドッキングするために必要なタンパクであるシナプトブレビンと GFP を組み合わせたレポーター UAS-nSyb::GFP を開発しました。このレポーターは、入力出力2種のシナプスのうち出力シナプスがある領域のみをラベルするので、神経細胞のどの部分から情報が送り出されるのかを知り、情報の流れを解析することができます。 さらに最近は、GABA レセプターに細胞タグを融合させたレポーターも開発されています。ショウジョウバエ脳では GABA レセプターは非常に多くの神経細胞に存在し、入力シナプスに局在していますので、nSyb::GFP と組み合わせて、細胞の入力出力の全貌が分かるわけです。 GFP 以外の蛍光タンパクGFP は青い光を当てると緑の蛍光を発し、FITC などの蛍光色素用に作られた従来の蛍光顕微鏡でそのまま観察できます。しかし二重染色のためには、他の色の蛍光タンパクも使えると便利です。GFP に変異を誘導して開発された、シアンの光を発する CFP や黄色い光を発する YFP は、明るさの点でも安定性の点でも、大変優れたレポーターです。しかし従来の蛍光顕微鏡のフィルターブロックや共焦点顕微鏡のレーザー光源がカバーしていない波長が励起に必要なため、実用上は多少不便な点があります。緑の光を当てると赤い光を発するような蛍光タンパクがあれば、ローダミンなどの蛍光色素用に作られた従来の蛍光顕微鏡でそのまま観察できるので、非常に便利です。サンゴからクローニングされた赤色蛍光タンパク DsRed は、この点では非常に好都合です。しかし改良が重ねられた GFP に比べると、初期の DsRed は暗くて使いにくいものでした。 多くの研究室がこれの改良に取り組みました。本研究室でも京大のグループと共同で、DsRed を改良して明るさを3倍に高めたレポーターを開発しました。GFP の場合、光量の増加は活性部位の構造の改良によって実現しましたが、DsRed ではそれよりも、フォールディングに影響する部分を改良し、翻訳されたもののうまくフォールディングされない分子の率を減らして収率を向上させたことが、光量増加の主な要因になっています。(最近では GFP でも、収率を向上させて光量を大きく増加させたものが広く利用されるようになりました。) DsRed は細胞全体を染め出すので、細胞の一部だけを染め出す GFP レポーターと組み合わせて用いるのと効果的です。たとえば出力シナプスを染め出す nSyb::GFP と DsRed を共発現させると、神経線維の中のどこに出力シナプスがあるかが詳しく解析できます。
GFP も DsRed も、mRNA から合成されたタンパクは時間をかけてフォールディングを行ない、最終的な形態に達して始めて蛍光を発します。GFP の場合は、この時間差は数十分ですが、DsRed では数時間~十数時間に達します。この時間差は一見すると欠点にしか見えませんが、工夫すれば非常に便利な使い方ができます。 UAS-GFP と UAS-DsRed を両方組み込んで、ひとつの細胞で同時に発現させます。発現が始まってから数時間は、GFP しか蛍光を発しませんが、時間が経つにつれて、DsRed の蛍光も観察されるようになります。このため、発現が始まってからの時間によって、細胞を違う色で染め分けることができるわけです。
隣接細胞をラベルする試み神経回路の結合様式を調べるには、GAL4 が発現する神経細胞だけでなく、その細胞とシナプス結合している細胞をラベルできれば大変便利です。この目的で開発されたのが、小麦胚芽由来のレクチンの一種である wheat germ agglutinin (WGA) を用いた UAS-WGA です。WGA は哺乳類の神経研究において、軸索に注入してそれと結合する隣接神経線維をラベルするのに昔から用いられてきたマーカーで、それを神経細胞内で合成させることで、隣接細胞をラベルしようと試みたわけです。 当初行なわれた実験では、一見シナプス特異的に隣接細胞をラベルしたように見える画像が論文に発表され、大きな注目を浴びました。しかしその後多くの研究者が試したところ、このようにシナプス特異的にラベルが伝わるように見えるのはごく一部の細胞だけであり、大多数のショウジョウバエ脳の神経細胞では、WGA はシナプスの有無に関係なく、近隣の細胞に非特異的に拡散してしまうことが分かりました。同様のことが線虫神経系でも明らかになっています。 レクチンは原理的に、細胞膜を自由に拡散して隣接細胞に伝達されうる分子であり、シナプス特異的に伝達されるような構造にはなっていません。しかし哺乳類の神経(特に長大な投射神経)では、神経線維の一本一本がグリアで覆われており、隣り合う神経細胞の膜同士が接しているのは、実質的にシナプスの部分だけです。そのため WGA は、シナプス特異的に伝わっているように見えるのだと思われます。線虫の神経系にはグリアはありませんし、ショウジョウバエの神経系ではグリアは神経線維の一本一本を包んでいるわけではありません。そのためレクチンははるかに容易に、シナプス以外の場所でも隣接細胞に伝わってしまうのだと思われます。真の意味でシナプス特異的に隣接細胞を染め出すようなレポーターは、現時点では残念ながら存在しません。 神経の活性を測定する試み(イメージングへの応用)多数の神経の活動をダイナミックに測定するために、細胞内部の電位や Ca 濃度、pH に応じて蛍光強度が変化する色素を使ったイメージングの手法が、1990年代に急速に発達しました。しかし Fura-2 のような染色液を標本に投与する従来の方法では、全ての神経細胞が一様にラベルされてしまい、測定しているのがどの種類の神経なのかが分かりません。 電位や Ca 濃度、pH などによって蛍光量が変化するタンパクの遺伝子を細胞特異的に発現させれば、特定の神経の活動だけをダイナミックに測定できます。pH に関しては、GFP を改変して pH による蛍光強度変化を大きくした変異タンパクを利用する方法が開発されています。 Ca 濃度に関しては、Ca 濃度によって構造が大きく変化するタンパクであるカルモジュリンを使った2つのアプローチが進められています。まず「カメレオン」は、CFP と YFP の間にカルモジュリンを挟んだ形のタンパクです。CFP しか励起されない波長の光を当てると、CFP が吸収したエネルギーが近くにある YFP に伝えられ、本来光らないはずの YFP からも蛍光が出るという FRET(蛍光エネルギー移動)という現象があります。FRET の量は CFP と YFP の間の距離によって大きく変わるので、Ca 濃度によってカルモジュリンの形状が変わると、蛍光強度が変化する仕組みです。また、「ペリカン」「カンガルー」「G-CaMP」などの蛍光タンパクは、GFP の途中にカルモジュリンを挟み込んだ形になっています。Ca 濃度によってカルモジュリンの形状が変わると、GFP の形状がゆがめられ、蛍光強度が変化する仕組みです。 脳の神経細胞で通常生じるレベルの Ca 濃度変動では G-CaMP が特に効果的で、多くの研究室で脳の様々な細胞における神経活動の測定が進められています。本研究室はそれ自身ではイメージングは行なっていませんが、これらの研究室と協力して、GAL4 エンハンサートラップ系統を用いて本研究室で同定した細胞で G-CaMP を発現させ、活性の解析を行なっています。 レポーター遺伝子の使い分けや、抗体染色や in situ 染色画像の解釈、分厚い標本の三次元再構成など、分子生物学的手法を用いた細胞のラベルには多くの注意しなくてはいけない点があります。見過ごされることが多いこれらのポイントを、以下の総説にまとめてあります。 Ito, K., Okada, R., Tanaka, N. K., and Awasaki, T. (2003). Cautionary observations on preparing and interpreting brain images using molecular biology-based staining techniques. Microsc Res Tech 62, 170-186. 2:ある遺伝子を発現する細胞の精密な同定
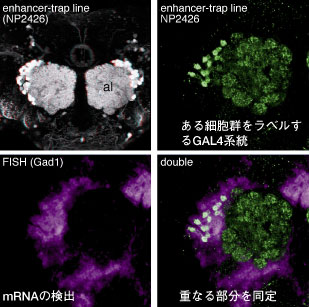
脳を4,000種類に染め分ける GAL4 エンハンサートラップ系統と、その画像データベースを利用することで、in situ 法で検出された脳領域にあるさまざまな細胞群を特異的にラベルするGAL4系統と組み合わせ、蛍光抗体染色で検出した GFP の画像と in situ で検出した mRNA の画像を対比し、 mRNA を発現する細胞を精密に同定することが可能になりました。 ゲノム研究が非常に進んでいるショウジョウバエでは、mRNA から作成された EST クローンのセットがストックセンターから分与されています。本研究室では、配布されている数千におよぶ EST クローンを自分の研究室で維持しています。自分が調べたい遺伝子をゲノムデータベースから探し、それに対応する EST クローンをフリーザーから取ってきて、GAL4 エンハンサートラップ系統と組み合わせて染めることで、様々な遺伝子の正確な発現パターンを極めて効率よく解析できるようになりました。 3:フリップアウト法を利用した単一細胞のラベル
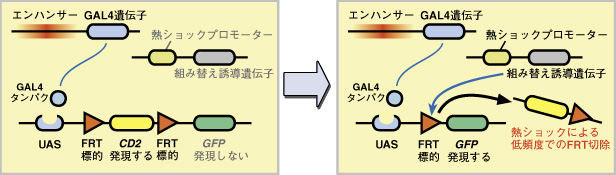
GAL4 エンハンサートラップ法では、同じエンハンサーが活性化されているすべての細胞で、GAL4 が発現します。これは、ある種類の神経細胞が作る回路全体の構造を知るのには便利ですが、細胞一つ一つが作る構造を調べることはできません。しかし GAL4 エンハンサートラップ法にさらにフリップアウト法を組み合わせることにより、GAL4 を発現する細胞のうちの1つだけの構造を取り出して可視化することができます。 まず、
このままの状態では、UAS によって発現が活性化されるのは CD2 遺伝子だけで、その後ろにあるレポーター遺伝 GFP は発現せず、細胞はラベルされません。発生の任意の時期に弱い熱ショックを加えると、組み替え誘導遺伝子 flippase がごく微量に発現します。flippase は2つのFRT 標的配列の間で組み替えを誘導し、間にあった CD2 遺伝子を除去します。しかし発現量が少ないので、この組み替えは低い頻度でしか起こりません。こうして、GAL4 を発現する細胞の中のごく一部だけで、GFP 遺伝子が発現するようになります。
4:FRT-GAL4法による、細胞系譜に依存したクローン構造のラベル
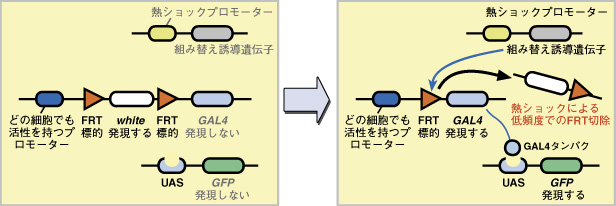
エンハンサーの活性パターンに応じて GAL4 を発現させるエンハンサートラップ法と全く異なり、本研究室で開発した FRT-GAL4 法は、細胞系譜に応じて特定の細胞に由来する子孫細胞だけで GAL4 を発現させるシステムです。 まず、
このままの状態では、アクチンプロモーターによって発現が駆動されるのは white 遺伝子だけで、GAL4 遺伝子は発現しないので、レポーター遺伝子 GFP も発現せず、細胞はラベルされません。 発生の任意の時期に弱い熱ショックを加えると、組み替え誘導遺伝子 flippase がごく微量に発現します。flippase は2つのFRT 標的配列の間で組み替えを誘導し、間にあった white 遺伝子を除去します。しかし発現量が少ないので、この組み替えは低い頻度でしか起こりません。 こうして、ごく一部の細胞だけで、white が除かれて GAL4 遺伝子が発現し、レポーター遺伝子 GFP も発現するようになります。これは、その細胞に由来する全ての子孫細胞にも伝えられ、細胞系譜に依存してクローン構造をラベルすることができます。
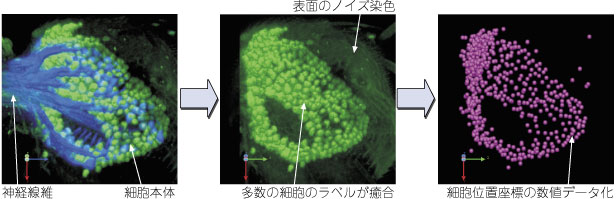
註:組み替えが幹細胞でなく分裂後の細胞で起こったばあいは、3:で紹介したフリップアウト法のように、単一細胞だけがラベルされます。 5:細胞位置情報の数値化 |






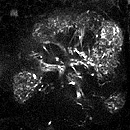
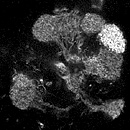

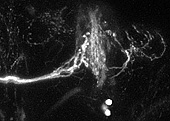
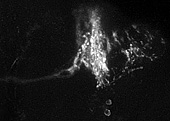
 ある遺伝子がどこで発現しているかを知るためには、メッセンジャー RNA の存在を検出する in situ ハイブリダイゼーション法が有効です。しかしメッセンジャー RNA は細胞体に集中して存在することが多いので、in situ 法では細胞体しかラベルできません。樹状突起や軸索など複雑な線維構造を持つ神経細胞では、細胞体だけ染まってもその神経の全体の形態が分からず、どんな細胞かを知ることができません。
ある遺伝子がどこで発現しているかを知るためには、メッセンジャー RNA の存在を検出する in situ ハイブリダイゼーション法が有効です。しかしメッセンジャー RNA は細胞体に集中して存在することが多いので、in situ 法では細胞体しかラベルできません。樹状突起や軸索など複雑な線維構造を持つ神経細胞では、細胞体だけ染まってもその神経の全体の形態が分からず、どんな細胞かを知ることができません。